ルイス式血液型
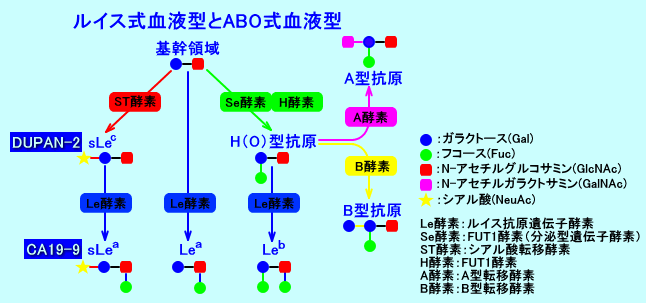
ルイス式血液型(Lewis式血液型)や、ABO式血液型の、血液型抗原は、タイプ1の基幹領域であるGalβ1→3GlcNAcに、糖転移酵素(Le酵素、Se酵素、ST酵素、A酵素、B酵素)により、糖(Gal、GlcNAc、GalNAc、Fuc、NeuAc)を結合して、合成される、糖鎖抗原。
Se酵素(FUT2)は、ガラクトース(Gal)に、フコース(Fuc)を、一個、α1→2結合させる。
Le酵素(FUT3)は、N-アセチルグルコサミン(GlcNAc)に、フコース(Fuc)を、一個、α1→4結合させる。FUT3(Le酵素)は、α1→3結合させる活性もある(α1,3 とα1,4の両方のフコース転移酵素活性がある)。
1.ルイス式血液型抗原の合成
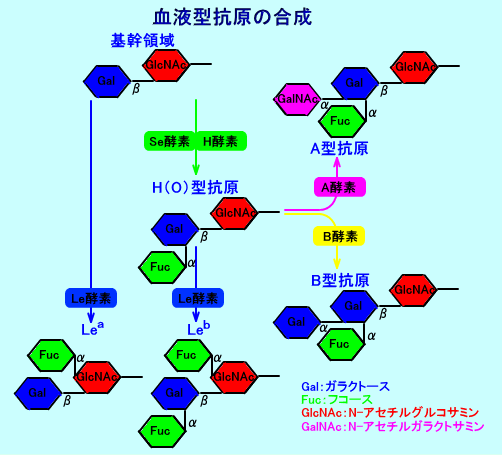
ルイス式血液型抗原は、ABO式血液型抗原(物質)と同一の前駆物質(タイプ1の基幹領域:Galβ1→3GlcNAc:一型糖鎖を構成)から、合成される。
ルイス式血液型抗原の合成には、フコース(Fuc)を結合させる二つの酵素、すなわち、Le酵素と、Se酵素(ABO式血液型物質の分泌液中への分泌を決定するに分泌型遺伝子酵素)が、関与する。
ルイス式血液型抗原は、基幹領域(タイプ1の基幹構造)のGalβ1→3GlcNAcから、Le酵素やSe酵素により、合成される。
上皮細胞で合成された、Lewis A抗原やLewis B抗原は、糖脂質として分泌され、赤血球に吸着する。
ルイス式血液型抗原は、成人では、赤血球以外にも、広く、多くの組織や臓器に、存在する:Lewis A抗原(Lea)は、膵臓、胆嚢、胃、大腸、の癌細胞や、少量だが、正常の上皮細胞に、存在する。
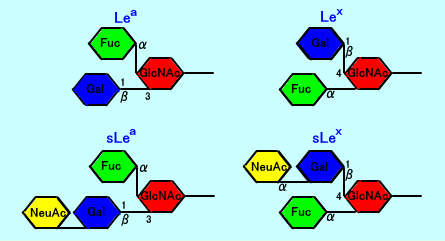
ルイス式血液型のLewis A抗原(Lea)は、Galβ1,3(Fucα1,4)GlcNAc-Rであり、フコース(Fuc)が、一個結合している。
Lewis A抗原(Lea)は、基幹領域(基幹構造)のGalβ1→3GlcNAc(注1)のN-アセチルグルコサミン(GlcNAc)に、Le酵素(ルイス抗原遺伝子酵素:FUT3酵素)により、フコース(Fuc)を、一個、1→4結合させ、合成される(注2)。なお、Le酵素(FUT3酵素:Fuc-TIII)は、フコース(Fuc)を、一個、1→4結合させる(α1,4 FUT)以外に、フコース(Fuc)を、一個、1→3結合させる(α1,3 FUT)作用もある。
b).Lewis B抗原(Leb)
ルイス式血液型のLewis B抗原(Leb)は、Fucα1,2Galβ1,3(Fucα1,4)GlcNAc-Rであり、フコース(Fuc)が2個結合している。
Lewis B抗原(Leb)の合成には、基幹領域のGalβ1→3GlcNAcのガラクトース(Gal)に、まず、Se酵素(分泌型遺伝子酵素:FUT2酵素)により、フコース(Fuc)を、1→2結合させ、H抗原(O型抗原)が形成される(注1)。次いで、基幹領域のGalβ1→3GlcNAcのN-アセチルグルコサミン(GlcNAc)に、Le酵素により、もうひとつフコース(Fuc)を、1→4結合させ、合成される。
上皮細胞で合成された、Lewis A抗原やLewis B抗原は、糖脂質として分泌され、赤血球に吸着する。
ルイス式血液型の赤血球の表現型は、4型に分類される:Le(a+b-)、Le(a-b+)、Le(a+b+)、Le(a-b-)。
日本人は、Le(a+b+)の人が多い:Le(a+b+) の人は、活性型のLe酵素と、活性がきわめて弱くなったSe酵素(Sej酵素)を持ち、部分的にフコース(Fuc)を結合させる。
Le(a-b+) の人は、 Se酵素、Le酵素ともに活性型であり、分泌型個体。Le(a-b+)の人は、Se酵素(分泌型遺伝子酵素)が存在する分泌型で、唾液中に、ABO式血液型のABH型物質が、認められる。
Le(a+b-)の人(ルイス抗原陽性の人)は、Se酵素を完全欠損していて、Lewis B抗原(Leb)を合成出来ない(Le酵素は、有している)。Le(a+b-)の人(Lewis抗原陽性の人)は、Se酵素(分泌型遺伝子酵素)が存在しない非分泌型で、唾液中に、ABO式血液型のABH型物質が、認められないか、ごく少量しか、認められない。Le(a+b-)の人(ルイス抗原陽性の人)は、非分泌型で、Lewis陽性ホモ(Le/Le, se/se)と、Lewis陽性ヘテロ(Le/le, se/se)とが、存在する。
Le(a-b-)の人(ルイス抗原陰性の人)は、Le酵素を欠損している(le/le)。Le(a-b-)の人には、Se酵素を有する分泌型の人と、Se酵素を有しない非分泌型の人とが、存在する。
表1 遺伝子とルイス式血液型の表現型 Le遺伝子 Se遺伝子 ABO式血液型物質の分泌 赤血球表面の表現型 Le/le Se/se 分泌 Le(a-b+) Le/Le Le/le Se/Se Le/Le Le/le se/se 非分泌 Le(a+b-) Le/Le le/le se/se 非分泌 Le(a-b-) Se/se 分泌 Se/Se
2.ABO式血液型抗原の合成
ABO式血液型物質の抗原決定基は、糖鎖であり、ルイス式血液型抗原と同一の前駆物質(タイプ1の基幹領域:Galβ1→3GlcNAc:一型糖鎖を構成)から、合成される。
基幹領域のGalβ1→3GlcNAc(一型糖鎖)のガラクトース(Gal)に、Se酵素(FUT2酵素:分泌型遺伝子酵素、α1,2 FUT)により、フコース(Fuc)を1→2結合させると、ABO式血液型(注2)の、H抗原(O型抗原:Fucα1→2Galβ1→3GlcNAc)が形成される。H抗原(O型抗原)は、A酵素やB酵素により、ABO式血液型物質(抗原)に合成されたり、Le酵素により、Lewis B抗原(Leb)に合成される。Se酵素(FUT2酵素)は、唾液中のH抗原(O型抗原)を合成するのに必要な酵素であり、Se酵素が欠損したホモ接合体は、非分泌型になる。
H抗原(O型抗原)は、H酵素(FUT1酵素、α1,2 FUT)によっても、一型糖鎖から合成される。H酵素(FUT1酵素:α1,2-fucosyltransferase:H遺伝子:H glycosyl transferase)は、赤血球上のABO式血液型物質(抗原)であるH抗原(O型抗原)を合成するのに、必要な酵素。H酵素(FUT1酵素)が変異する(劣性遺伝子hhのホモ結合体)と、ボンベイ型(bombay type)と言って、ABO式血液型物質(ABH式血液型物質)が、赤血球上に存在しないか、微量にしか存在しない為、A型、B型、AB型であっても、O型としか判定されない。ボンベイ型では、赤血球表面には、通常、存在するH抗原(O型抗原)が存在しないが、血清中には、抗H抗体が存在する(両親ともO型なの、A型やB型の子供が生まれることがある)。
フコース(Fuc)を1→2結合させる酵素(α1,2 FUT)であるSe酵素(FUT2酵素)や、H酵素(FUT1酵素)は、二型糖鎖(Galβ1→4GlcNAc)のガラクトース(Gal)に、フコース(Fuc)を1→2結合させることも可能。
H抗原(O型抗原)のGalに、A酵素(A型転移酵素:α-N-acetylgalactosaminyltransferase)により、N-アセチルガラクトサミン(GalNAc)が、α1→3結合すると、人間のABO式血液型の、A型抗原が形成される。
また、H抗原(O型抗原)のGalに、B酵素(B型転移酵素:galactosyltransferase)により、もう1つ、ガラクトース(Gal)Gal)が、α1→3結合すると、B型抗原が形成される。
糖が結合していない(0型)か、N-アセチルガラクトサミン(GalNAc)が結合する(A型)か、ガラクトース(Gal)が結合する(B型)か、わずか糖一つの違いが、ABO式血液型を決定している。
ABO式血液型抗原(ABO式血液型物質)は、赤血球で合成され、主に、赤血球表面に発現される。ABO式血液型抗原は、赤血球以外の血球細胞には、発現しない。
ABO式血液型抗原は、他の組織(消化管粘膜、腎臓、血管内膜、等)にも、発現される。
ABO式血液型では、ABO式血液型物質(ABH式血液型物質)を、体液中に多く分泌する分泌型と、少量しか分泌しない非分泌型(Se酵素が欠損したホモ接合体)とが、存在する。
日本人では、分泌型:非分泌型は、3:1の頻度で、存在する。
約77%の日本人は、分泌型(Se)で、赤血球の型と同じABO式血液型抗原(ABH型抗原物質)を分泌液中に含んでいる(唾液で判定する)。残り約23%の日本人は、非分泌型(se)。
Se酵素(FUT2酵素)は、分泌型遺伝子酵素:SeSe、Seseは、分泌型で、seseは、非分泌型。
分泌型の人では、唾液、精液に、ABO式血液型物質が、分泌され、検出される。
Le(a-b+) の人は、 Se酵素、Le酵素ともに活性型であり、分泌型になり、ABO式血液型抗原(ABH型抗原物質)と、ルイス式血液型のLewis B抗原(Leb)とを、唾液中などに、分泌している。
新生児では、赤血球膜上のA型抗原やB型抗原の量は、成人の30〜60%しか存在しない。成人では、赤血球膜上のA型抗原やB型抗原の量は、約80〜100万個/血球とされる。それに比して、臍帯血の赤血球では、赤血球膜上のA型抗原やB型抗原の量は、約20〜30万個/血球しか存在しない(成人の1/3〜1/4しか存在しない)。
新生児は、抗A抗体や抗B抗体の産生(腸内細菌の糖鎖に対して産生される)が不十分であり、また、母親の抗A抗体や抗B抗体が、胎盤を通過して、移行している。
その為、血液型の検査は、2歳以降に行う方が良い。3.シアリルLea抗原
シアリルLea抗原(シアリルルイスa抗原:sLea:CA19-9)の合成には、タイプ1の基幹領域のGalβ1→3GlcNAcのガラクトース(Gal)に、先ず、ST酵素(シアル酸転移酵素)により、シアル酸のN-アセチルノイラミン酸(NeuAc)を2→3結合させ、シアリルLec抗原(sLec:シアリルルイスc抗原:DUPAN-2)が形成される。
次いで、シアリルLec抗原(sLec)のGlcNAcに、Le酵素(ルイス抗原遺伝子酵素)により、フコース(Fuc)を1→4結合させて、シアリルLea抗原(CA19-9)が、合成される。
シアリルLea抗原は、結果的に、ルイス式血液型抗原のLewis A抗原(Lea)に、シアル酸(NeuAc)が、2→3結合した構造をしている。
シアリルLea糖鎖抗原(sLea:シアリルルイスa抗原)は、基幹領域がGalβ1→3GlcNAcで、ガラクトース(Gal)と、N-アセチルグルコサミン(GlcNAc)との結合が、1→3結合(Galβ1→3GlcNAc)をしている、一型糖鎖。
それに比して、シアリルLex鎖抗原(sLex:シアリルルイスX抗原)は、ガラクトース(Gal)と、N-アセチルグルコサミン(GlcNAc)との結合が、1→4結合(Galβ1→4GlcNAc)をしている、二型糖鎖(注3)。
4.Le(a+b-)とLe(a-b-)
ルイス式血液型は、Se酵素を持っていない人は、Le(a+b-)になり、Le酵素を持っていない人は、Le(a-b-)になる。
a).Le(a+b-)
ST酵素とSe酵素は、共存していると、基質(基幹領域)を競合する(ST酵素によりシアル酸を結合させsLecを経てsLeaを合成するか、Se酵素によりフコースを結合させLebを合成するか、競合する)。
Se酵素を持っていないLe(a+b-)の人(ルイスA抗原陽性の人)は、基質に、ST酵素によりシアル酸を結合させるので、シアリルLea糖鎖抗原の産生が増加して、癌がなくても、CA19-9値が高くなる。
b).Le(a-b-)
日本人では、人口の4〜10%の人は、Le酵素が欠損し、Le(a-b-)である(Lewis抗原陰性者)。
Le(a-b-)の人(ルイス抗原陰性者)は、シアリルLec抗原からシアリルLea抗原(sLea)を合成出来ない。
その結果、Le酵素が欠損しているLe(a-b-)の人は、癌があっても、CA19-9値(シアリルLea抗原)は、上昇しない(ゼロになる)。
しかし、Le(a-b-)の人は、癌があれば、シアリルLec抗原(sLec)は、上昇し得るので、DUPAN-2の測定が有用である。DUPAN-2(シアリルLec抗原:sLec)は、CA19-9(シアリルLea抗原:sLea)の前駆体である(上図)。
なお、CA50は、シアリルLec抗原も検出するので、Le(a-b-)の人の診断に利点があると言われて来たが、CA50は、シアリルLea抗原をより多く検出する。
また、SLX(シアリルLex抗原)の合成に際して、N-アセチルグルコサミン(GlcNAc)に、フコース(Fuc)を結合させるには、Le酵素(FUT3酵素:α1.3とα1.4の両方のフコース転移酵素活性を示す)と、FUT6酵素(α1.3のフコース転移酵素活性を示す)が用いられる。SLX(シアリルLex抗原)のフコースは、N-アセチルグルコサミン(GlcNAc)に、α1.3結合(1→3結合)している。そのため、Le酵素が欠損しているLe(a-b-)の人でも、FUT6酵素により、SLX(シアリルLex抗原)は合成される。その結果、Le(a-b-)の人でも、癌があると、SLX値は上昇するので、Le(a-b-)の人では、CA19-9より、SLXを測定する。
4.シスAB型
ABO血液型遺伝子は、ヒトでは、9番染色体に存在する。
ABO式血液型を決定する、A型遺伝子(A酵素によりA型抗原を生成する)や、B型遺伝子(B酵素によりB型抗原を生成する)は、9番染色体の長腕(9q34)位置に存在する。
ABO式血液型がAB型の人は、二対ある9番染色体の内、片方は、A型遺伝子で、他方は、B型遺伝子になっている。
シスAB型(cis-AB型)では、変異したA型遺伝子(AB型遺伝子)が存在し、A型抗原を生成するのみならず(GalNAcを結合させる)、B型抗原を生成する(Galを結合させる)。通常のAB型の人は、9番染色体の片方がA型遺伝子で、他方はB型遺伝子。
シスAB型の人は、10万人に1人の割合で存在する。
通常は、父親がAB型で、母親がB型の両親からは、O型の子供は生まれない。
しかし、父親がシスAB型(9番染色体の片方がcis-AB型遺伝子で、他方がO型遺伝子)だと、母親がB型(9番染色体の片方がB型遺伝子で、他方がO型遺伝子)であっても、O型の子供が生まれることが有り得る。
注1:GalとGlcNAcとが結合した2糖構造は、色々な糖鎖の基幹領域(基幹構造)になっている。
基幹領域には、タイプ1(Galβ1→3GlcNAc:一型糖鎖を構成)と、タイプ2(Galβ1→4GlcNAc:二型糖鎖を構成)とが、存在する。タイプ2の基幹領域(Galβ1→4GlcNAc)は、N-アセチルラクトサミン(N-acetyllactosamine:LacNAc)。
ABO式血液型物質や、ルイス式血液型抗原や、シアリルLea糖鎖抗原(sLea:CA19-9)は、タイプ1の基幹領域(Galβ1→3GlcNAc:一型糖鎖を構成)から構成される。
それに比して、シアリルLex鎖抗原(sLex:SLX)は、タイプ2の基幹領域(Galβ1→4GlcNAc:二型糖鎖を構成)から構成されている。
注2:ABO式血液型抗原は、赤血球だけでなく、腸の粘膜上皮細胞の表面にも、存在する。
ABO式血液型抗原は、腸では、糖鎖として、腸内細菌との結合能に、影響する。例えば、A型の人の腸内では、納豆菌が定着しやすいという。
血清中に、A型の人は、B型の赤血球を凝集させる抗B抗体(β:IgMクラス)を有している。この抗B抗体は、自分の体内に侵入した微生物のB型糖鎖に感作され、産生される。
ニホンザルは、B型しか存在しない(ニホンザルは、B型抗原しか作れず、A型抗原は作れない)。
なお、ABO式血液型抗原は、一つの遺伝子に乗って存在している。血液型A型の人は、遺伝子型レベルでは、AA型の人と、AO型の人とがいる。
稀に、同一染色体上に、AとBの遺伝子が乗る場合(cis AB)があり、その場合、片親がAB型なのに、O型の子供が生まれることがある(逆に、片親がO型なのに、AB型の子供が生まれることがある)。
Lewis抗原やシアリルLex鎖抗原(sLex:SLX)などは、上皮細胞で産生される。ABO血液型物質(ABO式血液型抗原)は、赤血球で合成される。
唾液中のH抗原(O型抗原)は、Se酵素(FUT2酵素)により合成され、赤血球のH抗原(O型抗原)は、H酵素(FUT1酵素)により合成される。
H抗原(O型抗原)は、基幹領域のGalβ1→3GlcNAc(一型糖鎖)のガラクトース(Gal)に、Se酵素(FUT2酵素:分泌型遺伝子酵素:上皮細胞)か、H酵素(FUT1酵素:赤血球など)により、フコース(Fuc)が、1→2結合して、生成される。
Se酵素(FUT2酵素)やH酵素(FUT1酵素)は、α1,2 FUT。
H酵素(FUT1酵素)とSe酵素(FUT2酵素)の組織分布は、解明されていない。胃や大腸などの消化管組織の上皮細胞には、H酵素とSe酵素の両方が、発現していると言う。
注3:Lex抗原(Lewis X抗原:SSEA-1抗原)は、Lea抗原(Lewis A)と相似して、フコース(Fuc)が、N-アセチルグルコサミン(GlcNAc)に結合している。しかし、Lea抗原では、基幹構造が、Galβ1→3GlcNAc(タイプ1)なのに対して、Lex抗原では、Galβ1→4GlcNAc(タイプ2)。
Lex抗原:Galβ1,4(Fucα1,3)GlcNAc-R
Lea抗原:Galβ1,3(Fucα1,4)GlcNAc-R
Lex抗原に、ST酵素により、シアル酸(NeuAc)が結合した糖鎖が、sLex抗原(シアリルLex抗原)。
参考文献
・腫瘍マーカー―その診断的意義と今後の展望 日本医師会雑誌 第131巻・第5号 2004年.
・渡邊弘之、澤武紀雄:CA19-9とCA50、日本医師会雑誌 第131巻・第5号 、619-623頁、2004年(平成16年).
・小田桐恵美:CA19-9 内科 61巻6号、1386-1387、1988年(南江堂).
・上田英雄、他:内科学(第四版、1987年、朝倉書店).
・Maciej Ugorski, et al: Sialyl Lewisa: a tumor-associated carbohydrate antigen involved in adhesion and metastatic potenrial of cancer cells. Acta Biochimica Polonica Vol.49 No.2/2002, 303-311.
・Meike Rinnbauer, et al: Epitope mapping of sialyl Lewisx bound to E-selectin using saturation transfer difference NMR experiments. Glycobiology, 2003, Vol. 13, No. 6 435-443.
・浅井隆善:造血幹細胞移植とABO血液型(質疑応答) 日本醫事新報 No.4136(2003年8月2日)、90-91頁.
・古川研:血液型の遺伝 日本醫事新報 No.2994(昭和56.9.12日)、166-167.