丂
丂 丂
丂 丂
丂
丂丂丂
丂MHC
丂MHC偲偼丄major histocompatibility complex偺棯偱丄庡梫慻怐揔崌峈尨暋崌懱偺偙偲丅
丂僸僩偺MHC偼丄HLA乮human leukocyte antigen乯偲屇偽傟丄僸僩戞6愼怓懱抁榬忋偵丄堚揱巕椞堟偑懚嵼偡傞丅
丂儅僂僗偺MHC偼丄H-2峈尨偲屇偽傟傞丅
丂峈尨採帵嵶朎昞柺偺NHC暘巕偼丄T嵶朎偵丄峈尨傪採帵偡傞乮峈尨採帵暘巕乯丅
丂僿儖僷乕T嵶朎偼丄庡偵丄MHC僋儔僗I俬暘巕偵寢崌偟偨嵶朎奜峈尨乮嵶嬠傗僂僀儖僗桼棃偺抈敀側偳乯偵斀墳偟丄僉儔乕T嵶朎偼丄庡偵丄MHC僋儔僗I暘巕偵寢崌偟偨嵶朎撪峈尨乮庮釃摿堎峈尨側偳乯偵斀墳偡傞丅
丂MHC乮HLA乯偼丄憻婍堏怉偺慻怐揔崌惈峈尨丄幘姵偺敪徢偲娭楢偡傞幘姵姶庴惈峈尨丄敀寣媴丒寣彫斅偺桝寣暃嶌梡偵娭學偡傞峈尨丄恊巕娪掕偺儅乕僇乕偲偟偰丄椪彴揑偵丄廳梫丅
丂侾丏MHC僋儔僗I暘巕偲丄MHC僋儔僗II暘巕
丂MHC偼丄MHC僋儔僗俬暘巕偲MHC僋儔僗I俬暘巕偺俀庬椶懚嵼偡傞丅
丂MHC暘巕偼丄嵶朎撪偱惗惉偝傟丄嵶朎昞柺偵桝憲偝傟傞丅
丂MHC暘巕偼丄嵟傕奜懁偺晹暘偵丄俀杮偺兛嵔乮兛傊儕僢僋僗乯偵傛傝峚偑宍惉偝傟偰偄傞丅峈尨採帵嵶朎側偳偺嵶朎撪偱丄惗惉偝傟偨MHC峚偵丄峈尨儁僾僠僪偑寢崌偟丄MHC亄峈尨儁僾僠僪偺暋崌懱偲偟偰丄嵶朎枌昞柺偵桝憲偝傟丄敪尰偡傞丅
丂T嵶朎乮僿儖僷乕T嵶朎丄僉儔乕T嵶朎乯偼丄嵶朎昞柺偵懚嵼偡傞T嵶朎峈尨庴梕懱乮TCR乯偵傛傝丄峈尨採帵嵶朎乮儅僋儘僼傽乕僕側偳乯傗丄昗揑嵶朎偺昞柺偵懚嵼偡傞MHC亄峈尨儁僾僠僪偺暋崌懱傪丄擣幆偡傞丅
丂侾乯丏MHC僋儔僗俬暘巕
丂MHC僋儔僗俬暘巕偼丄兛嵔偲丄兝2儈僋儘僌儘僽儕儞偺兝嵔偱峔惉偝傟傞丅
丂僸僩偱偼丄MHC僋儔僗俬暘巕乮MHC僋儔僗俬峈尨乯偲偟偰丄HLA-A丄HLA-B丄HLA-C偑懚嵼偡傞丅
丂MHC僋儔僗俬暘巕偼丄T嵶朎丄B嵶朎偺懠丄傎偲傫偳慡偰偺嵶朎昞柺傗丄寣彫斅昞柺偵丄敪尰偟偰偄傞丅
丂嵶朎偵姶愼偟偨僂僀儖僗偼丄嵶朎幙撪偱僂僀儖僗抈敀幙傪崌惉偡傞丅崌惉偝傟偨僂僀儖僗抈敀幙乮嵶朎撪峈尨乯偼丄僾儘僥傾僜乕儉偱丄峈尨儁僾僠僪乮傾儈僲巁偺悢偑俈乣俋働乯偵暘夝偝傟丄慹柺彫朎懱乮ER乯偵塣偽傟偰丄崌惉搑拞偺MHC僋儔僗俬暘巕偲寢崌偟偰丄MHC僋儔僗俬暘巕亄峈尨儁僾僠僪偲偟偰丄嵶朎昞柺偵敪尰偝傟傞丅
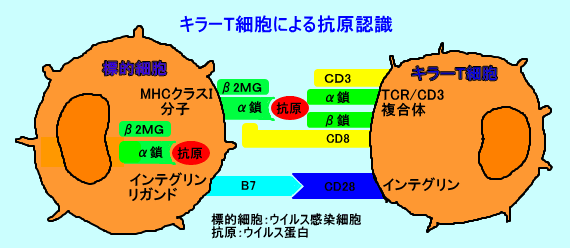
丂偙偺傛偆偵偟偰丄昗揑嵶朎乮庮釃嵶朎丄僂僀儖僗姶愼嵶朎乯昞柺偺MHC僋儔僗俬暘巕偼丄嵶朎撪偱崌惉偝傟偨峈尨儁僾僠僪乮嵶朎撪峈尨乯傪寢崌偝偣傞丅偦偟偰丄昗揑嵶朎偼丄僂僀儖僗側偳偺峈尨傪丄MHC僋儔僗俬暘巕亄峈尨儁僾僠僪偲偟偰丄CD8梲惈僉儔乕T嵶朎昞柺偺T嵶朎峈尨庴梕懱乮TCR乯偵丄採帵偡傞丅
丂CD8梲惈僉儔乕T嵶朎偼丄僂僀儖僗峈尨傪敪尰偟丄帺屓偲摨偠MHC僋儔僗俬暘巕傪敪尰偟偨丄僂僀儖僗姶愼嵶朎傪丄忈奞偡傞乮嶦偡乯丅偟偐偟丄CD8梲惈僉儔乕T嵶朎偼丄僂僀儖僗偦偺傕偺傪丄嶦偡偺偱偼側偄乮CD8梲惈僉儔乕T嵶朎偼丄僂僀儖僗姶愼嵶朎傪嶦偡偙偲偱丄僂僀儖僗偺憹怋傪梷惂偡傞乯丅
丂MHC僋儔僗II暘巕偼丄兛嵔乮兛1丄兛2乯偲丄兝嵔乮兝1丄兝2乯偱峔惉偝傟傞丅
丂僸僩偱偼丄MHC僋儔僗俬I暘巕偲偟偰丄HLA-DR丄HLA-DP丄HLA-DQ偑懚嵼偡傞丅
丂MHC僋儔僗俬I暘巕乮MHC僋儔僗俬I峈尨乯偼丄Ia峈尨偲傕屇偽傟偨丅
丂MHC僋儔僗俬俬暘巕偼丄尷傜傟偨嵶朎偺昞柺偵敪尰偡傞丗庽忬嵶朎乮旂晢儔儞僎儖僴儞僗嵶朎側偳乯丄扨媴丒儅僋儘僼傽乕僕丄B嵶朎丄妶惈壔T嵶朎乮惷巭婜偺T嵶朎偼丄敪尰偟偰偄側偄乯丄嫻態忋旂嵶朎丄惛巕側偳丅
丂僋儔僗俬俬峈尨偺愭抂晹暘偵偼丄峈尨儁僾僠僪偲寢崌偡傞偨傔偺丄峚乮antigen binding groove丗峈尨寢崌鈢丄binding cleft乯偑懚嵼偡傞丅
丂嵶嬠偼丄僄儞僪僒僀僩乕僔僗偱嵶朎撪偵庢傝崬傑傟偰丄儕僜僜乕儉偱峈尨儁僾僠僪偵暘夝偝傟丄MHC僋儔僗II暘巕偺峚偵寢崌偟偰丄嵶朎昞柺偵敪尰偝傟傞丅
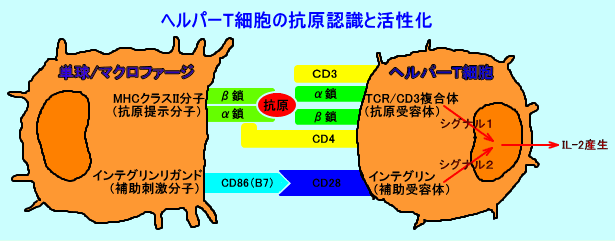
丂偙偺傛偆偵偟偰丄峈尨採帵嵶朎乮儅僋儘僼傽乕僕側偳乯昞柺偺MHC僋儔僗II暘巕偼丄奜棃惈抈敀乮嵶朎奜峈尨乯偺峈尨儁僾僠僪乮惛惢僣儀儖僋儕儞側偳丄拲侾乯傪丄polyproline type II偲偄偆堷偒墑偽偝傟偨宍偱丄MHC僋儔僗II暘巕偺峚偵寢崌偝偣丄CD4梲惈僿儖僷乕T嵶朎昞柺偺T嵶朎峈尨庴梕懱乮TCR乯偵採帵偡傞丅
丂MHC僋儔僗II暘巕傪峔惉偡傞傾儈僲巁巆婎偑丄MHC乮HLA乯偺宆偵傛偭偰丄堚揱揑偵堎側傞偨傔丄MHC乮HLA乯偺宆偵傛偭偰丄MHC僋儔僗II暘巕偺峚偼丄堎側傞丅偦偺偨傔丄摨偠峈尨儁僾僠僪偱偁偭偰傕丄MHC僋儔僗II暘巕偺峚傊偺寢崌偟堈偝偼丄MHC乮HLA乯偺宆偵傛偭偰丄堎側傞丅
丂偦偺堊丄MHC乮HLA乯偺宆偵傛偭偰丄峈尨乮峈尨儁僾僠僪乯偵懳偡傞柶塽墳摎偺嫮搙偑丄堎側傞丅
丂T嵶朎乮僿儖僷乕T嵶朎丄僉儔乕T嵶朎乯偼丄T嵶朎峈尨庴梕懱乮T cell receptor丗TCR乯傪峈尨庴梕懱偲偟偰丄摿堎峈尨乮峈尨儁僾僠僪乯偲斀墳偡傞丅
丂T嵶朎峈尨庴梕懱乮TCR乯偼丄CD3暘巕偲丄嵶朎枌娧捠晹偱丄旕嫟桳揑偵寢崌偟丄TCR丒CD3暋崌懱乮兛兝/兞兠/兟兠/兡-兡乯傪丄宍惉偟偰偄傞丅
丂B嵶朎偼丄嵶朎枌昞柺偺枌宆柶塽僌儘僽儕儞乮immunoglobulin丗Ig乯傪峈尨庴梕懱偲偟偰丄摿堎峈尨乮峈尨儁僾僠僪乯偲斀墳偡傞丅
丂T嵶朎偱偼丄T嵶朎峈尨庴梕懱乮TCR乯偼丄MHC乮庡梫慻怐揔崌峈尨暋崌懱乯偲峈尨儁僾僠僪偲偺暋崌懱乮MHC亄峈尨儁僾僠僪乯偵丄斀墳偡傞丅
丂僿儖僷乕T嵶朎乮CD4梲惈T嵶朎乯偺T嵶朎峈尨庴梕懱乮TCR乯偼丄峈尨儁僾僠僪傪寢崌偝偣偨MHC僋儔僗II暘巕偵斀墳偟丄僉儔乕T嵶朎乮CD8梲惈T嵶朎乯偺T嵶朎峈尨庴梕懱乮TCR乯偼丄峈尨儁僾僠僪傪寢崌偝偣偨MHC僋儔僗I暘巕偵斀墳偡傞丅峈尨採帵嵶朎乮儅僋儘僼傽乕僕側偳乯偺MHC僋儔僗II暘巕偼丄庡偵丄嵶朎奜偐傜婷怘偟偨峈尨儁僾僠僪傪寢崌偝偣丄僿儖僷乕T嵶朎偵丄峈尨偲偟偰採帵偡傞丅傑偨丄庮釃嵶朎側偳偺MHC僋儔僗II暘巕偼丄庡偵丄嵶朎撪峈尨傪峈尨儁僾僠僪偲偟偰寢崌偝偣丄僉儔乕T嵶朎偵丄峈尨偲偟偰採帵偡傞丅
丂廬偭偰丄僿儖僷乕T嵶朎偼丄庡偵丄嵶朎奜峈尨乮嵶嬠傗僂僀儖僗桼棃偺抈敀乯偵斀墳偟丄傑偨丄僉儔乕T嵶朎偼丄庡偵丄嵶朎撪峈尨乮庮釃摿堎峈尨丄僂僀儖僗偑敪尰偝偣傞抈敀乯偵斀墳偡傞丅
丂B嵶朎偱偼丄枌宆柶塽僌儘僽儕儞乮Ig乯偼丄峈尨儁僾僠僪偦偺傕偺偵丄斀墳偡傞丅
丂俁丏MHC偵傛傞帺屓擣幆
丂柶塽宯偱傕丄T嵶朎偼丄帺屓偺MHC乮HLA乯偑曄壔乮曄惈乯偟偰偄側偄偐丄擣幆偡傞丅T嵶朎偼丄TCR乮T嵶朎庴梕懱乯偵傛傝丄峈尨採帵嵶朎昞柺偺丄宆偺堎側傞MHC乮HLA乯傗丄曄惈偟偨MHC乮HLA乯傪丄乽旕帺屓乿偲偟偰擣幆偡傞偲峫偊傜傟傞丅
丂TCR乮T嵶朎庴梕懱乯偼丄MHC乮HLA乯偲峈尨儁僾僠僪偺暋崌懱乮MHC亄峈尨儁僾僠僪丗MHC偲丄偦偺峚偵寢崌偟偨峈尨儁僾僠僪乯傪擣幆偡傞丅
丂侾乯丏嵶朎昞柺偺MHC僋儔僗I暘巕乮兛嵔乯偼丄NK嵶朎偵丄帺屓偱偁傞偙偲傪丄擣幆偝偣傞
丂尨巒揑側妉摼柶塽偱偁傞丄NK嵶朎偼丄NK慾巭儗僙僾僞乕乮MHC僋儔僗I暘巕庴梕懱乯偵傛傝丄昗揑嵶朎昞柺偵丄帺屓偲摨偠MHC僋儔僗I暘巕乮兛嵔乯傪擣幆偡傞偲丄帺屓乮枴曽乯偲擣幆偟丄峌寕偟側偄丅偦偟偰丄NK嵶朎偼丄昗揑嵶朎昞柺偵丄帺屓偲摨偠MHC僋儔僗I暘巕乮兛嵔乯傪擣幆弌棃側偄偲丄峌寕偡傞丅尵傢偽丄愴応偱偼丄枴曽乮帺屓乯偩偲偄偆栦復傪偮偗偰偄側偄暫傪丄揋偲尒側偡傛偆偵丄NK嵶朎偼丄帺屓偱偁傞偲徹柧偡傞MHC僋儔僗I暘巕乮兛嵔乯傪丄敪尰偟偰偄側偄嵶朎傪丄旕帺屓乮揋乯偲丄尒側偡庤朄偩丅偮傑傝丄NK嵶朎偼丄帺屓乮枴曽乯偱偁傞徹柧偑偁傞偐丄娔帇偟偰偄傞偲丄峫偊傜傟傞丅
丂偙偺傛偆偵丄昗揑嵶朎偺嵶朎昞柺偺MHC僋儔僗I暘巕乮兛嵔乯偼丄NK嵶朎偵丄帺屓偱偁傞偙偲傪丄擣幆偝偣傞丅
丂俀乯丏嵶朎昞柺偺MHC僋儔僗俬俬暘巕偼丄峈尨儁僾僠僪傪寢崌偝偣丄僿儖僷乕T嵶朎偵丄旕帺屓偱偁傞偙偲傪丄擣幆偝偣傞
丂懠曽丄僿儖僷乕T嵶朎偺條偵丄T嵶朎偼丄TCR乮T嵶朎庴梕懱乯偱丄扨媴丒儅僋儘僼傽乕僕昞柺偺丄MHC僋儔僗俬俬暘巕偲峈尨儁僾僠僪偺暋崌懱傪擣幆偡傞丅偙傟偼丄偳偪傜偐偲尵偊偽丄帺屓偺MHC乮HLA乯偑曄惈偟偰偄側偄偐丄擣幆偟丄曄惈偟偰偄側偗傟偽帺屓乮枴曽乯丄曄惈偟偰偄傟偽旕帺屓乮揋乯偲丄擣幆偡傞丅偮傑傝丄T嵶朎偼丄帺屓偑曄惈偟偰偄側偄偐丄娔帇偟偰偄傞偲丄峫偊傜傟傞丅偙傟偼丄尵傢偽丄愴応偱偼丄枴曽乮帺屓乯偑丄棤愗偭偰偄側偄偐乮曄惈偟偰偄側偄偐乯丄娔帇偟偰丄揋偺栦復傪晅偗偰偄側偗傟偽枴曽乮帺屓乯丄揋偺栦復傪晅偗偰偄傟偽揋乮旕帺屓乯偲丄尒側偡傛偆側庤朄偩丅
丂偙偺傛偆偵丄峈尨採帵嵶朎乮儅僋儘僼傽乕僕側偳乯偺嵶朎昞柺偺MHC僋儔僗俬俬暘巕偼丄峈尨儁僾僠僪傪寢崌偝偣丄僿儖僷乕T嵶朎偵丄旕帺屓乮曄惈偟偨帺屓乯偱偁傞偙偲傪丄擣幆偝偣傞丅
丂側偍丄昗揑嵶朎乮僂僀儖僗姶愼嵶朎乯偺嵶朎昞柺偺MHC僋儔僗俬俬暘巕偼丄峈尨儁僾僠僪傪寢崌偝偣丄僉儔乕T嵶朎偵丄旕帺屓乮曄惈偟偨帺屓乯偱偁傞偙偲傪丄擣幆偝偣傞丅
丂俁乯丏堏怉曅偺MHC暘巕偼丄旕帺屓偲娕橍偝傟丄堏怉嫅愨斀墳偑婲偙傞
丂堏怉儗僔僺僄儞僩偺堏怋偟偨慻怐偵丄嫅愨斀墳偑婲偙傞帪丄僪僫乕桼棃偺堏怉曅偺嵶朎昞柺偺傾儘MHC乮allo MHC乯偼丄旕帺屓偲偟偰擣幆偝傟偰偄傞丅
丂僪僫乕偺傾儘MHC乮摨庬堎宯HLA乯偑丄旕帺屓偲擣幆偝傟傞偺偼丄堏怉儗僔僺僄儞僩偺T嵶朎偑丄嵶朎昞柺偺TCR乮T嵶朎庴梕懱乯偵傛傝丄帺屓偺MHC偲堎側傞丄僪僫乕偺傾儘MHC乮摨庬堎宯HLA乯傪丄帺屓MHC亄峈尨儁僾僠僪偲丄擣幆偡傞堊偲丄巚傢傟傞丅
丂MHC僋儔僗俬I暘巕偺HLA-DR乮DRB1乯偑堎側傞丄擇恖偺僸僩偺儕儞僷媴傪丄帋尡娗撪偱丄崿偤偰攟梴偡傞乮崿崌攟梴偡傞乯偲丄T嵶朎乮T儕儞僷媴乯偑丄屳偄偵丄懠偺僸僩偺儕儞僷媴偺HLA-DR暘巕傪丄傾儘峈尨偲偟偰擣幆偟丄妶惈壔偝傟丄憹怋斀墳傪婲偙偡乮儕儞僷媴崿崌斀墳丗mixed lymphocyte reaction丗MLR乯丅
丂MLR偵嵺偟偰偼丄T嵶朎偺撪丄悢乣悢廫亾偺嵶朎偑丄妶惈壔偝傟丄憹怋斀墳傪婲偙偡丅
丂係丏MHC偺宆偵傛傝丄摿掕偺姶愼徢偵滊偭偨帪偺丄徢忬偺嫮偝偑丄堎側傞
丂MHC乮HLA乯偺宆偵傛傝丄摿掕偺峈尨偵懳偡傞丄柶塽墳摎偺嫮搙偑堎側傞丅
丂偙傟偼丄摨偠峈尨儁僾僠僪偱偁偭偰傕丄MHC乮HLA乯偺宆偵傛傝丄MHC乮HLA乯偺峚傊寢崌偟堈偐偭偨傝丄寢崌偟偵偔偄偨傔偲偝傟傞丅
丂椺偊偽丄僸僩偺HLA偺DR51-B5偼丄僀儞僼儖僄儞僓僂僀儖僗偺HA偲丄嫮偔寢崌偡傞丅
丂僸僩偺HLA偺DQ6-B5偼丄梟寣惈楢嵔媴嬠乮梟楢嬠乯偺M抈敀偲丄嫮偔寢崌偡傞丅
丂廬偭偰丄HLA偺DR51-B5傪桳偡傞僸僩偼丄僀儞僼儖僄儞僓僂僀儖僗偵懳偟偰丄傑偨丄HLA偺DQ6-B5傪桳偡傞僸僩偼丄梟楢嬠偵懳偟偰丄嫮偄柶塽墳摎傪帵偡丅
丂偙偺傛偆偵丄MHC乮HLA乯偺宆偵傛傝丄摿掕乮栤戣乯偺峈尨偲寢崌偺偟堈偝偵嵎偑偁傞丅
丂MHC乮HLA乯偺宆偵傛傝丄摿掕偺峈尨偵懳偟偰丄柶塽墳摎偡傞偐丄柶塽旕墳摎側偺偐丄寛傑傞丅
丂廬偭偰丄摨偠姶愼徢偵滊偭偨応崌偱傕丄屄懱偺MHC乮HLA乯偺宆偵傛傝丄徢忬偺嫮偝丄昦忬偺宱夁側偳偑丄堎側傞丅
丂偙偺偙偲偼丄摨偠庬偑丄堎側傞乮懡條惈偺偁傞乯MHC乮HLA乯傪桳偡傞偙偲偵傛傝丄懡庬偺姶愼徢偵滊偭偨応崌丄偳傟偐偺屄懱偑丄惗偒墑傃傞堊偵丄搒崌偑椙偄丅
丂柶塽墳摎偑嫮偡偓傞乮崅偡偓傞乯偙偲偼丄壴暡徢側偳丄傾儗儖僊乕幘姵側偳傪棃偨偟丄惗懱偵晄棙側応崌傕偁傞丅
丂俆丏CD4梲惈偺傾儘MHC峈尨斀墳惈僉儔乕T嵶朎
丂堦斒揑偵偼丄CD4梲惈嵶朎偼僿儖僷乕T嵶朎丄CD8梲惈嵶朎偼僉儔乕T嵶朎偲丄暘椶偝傟傞丅
丂偟偐偟丄CD4梲惈嵶朎偵偼丄傾儘峈尨偵懳偟偰丄僉儔乕T嵶朎妶惈乮嵶朎忈奞妶惈乯傪桳偡傞嵶朎傕丄懚嵼偡傞丅椺偊偽丄揱愼惈扨妀姵帣偺枛徑寣扨妀媴乮CD8梲惈嵶朎偑憹壛偟偰偄傞丗CD4/CD8亖8/66乯傪丄EBV乮Epstein-Barr virus乯偱宍幙揮姺乮transform乯偟偨丄懠恖桼棃乮傾儘桼棃乯偺B嵶朎姅乮B儕儞僷夎媴丗LCL乯偱巋寖偟側偑傜丄recombinant IL-2傪1U/ml揧壛偟偰丄挿婜攟梴偡傞偲丄CD4梲惈丄HLA-DR梲惈乮MHC僋儔僗II暘巕乯梲惈偺T嵶朎姅偑丄庽棫偝傟傞丅丂
丂偙偺挿婜攟梴偝傟偨丄CD4梲惈T嵶朎姅偼丄巋寖偵梡偄偨傾儘桼棃偺B嵶朎姅乮allogenic LCL-MY乯傗丄PHA偱巋寖偟偨傾儘桼棃偺儕儞僷夎媴偵懳偟偰丄嫮偄嵶朎忈奞妶惈傪帵偡偑丄帺屓桼棃偺B嵶朎姅乮autologous LCL-YT乯偵懳偟偰偼丄嵶朎忈奞妶惈傪帵偝側偄丅
丂廬偭偰丄偙偺傛偆側丄CD4梲惈T嵶朎姅偼丄傾儘峈尨乮allo MHC乯偵懳偟偡傞僉儔乕妶惈乮嵶朎忈奞妶惈乯傪桳偡傞丄傾儘MHC峈尨斀墳惈僉儔乕T嵶朎乮allo MHC-reactive cytotoxic T lymphocytes乯 偲丄峫偊傜傟傞丅丂
丂
丂僗僥儘僀僪嵻乮僐僴僋巁僾儗僪僯僝儘儞僫僩儕僂儉乯傪丄recombinant IL-2乮1U/ml乯傪梡偄偰丄挿婜攟梴拞偺CD4梲惈T嵶朎姅乮T儕儞僷媴乯偵揧壛偡傞偲丄惗嵶朎悢偲丄昗揑嵶朎忈奞妶惈偑丄尭彮偡傞丗僗僥儘僀僪嵻偑丄10-8乣10-9M偺擹搙偱偼丄惗嵶朎悢偼桳堄偵尭彮偟側偄乮T儕儞僷媴偺憹怋傪丄桳堄偵梷惂偟側偄乯丅偟偐偟丄昗揑嵶朎忈奞妶惈偼丄10-8乣10-9M偺擹搙偱傕丄梷惂偝傟傞乮10-9M偑丄惗棟揑側慻怐拞擹搙偲尵傢傟傞乯丅
丂拲侾丗寢妀梊杊朄偑夵惓偝傟丄暯惉17擭4寧偐傜丄僣儀儖僋儕儞斀墳傪偟側偄偱丄BCG愙庬偑峴傢傟傞傛偆偵側偭偨丅
丂傕偟丄婛偵寢妀嬠偵姶愼偟偰偄傞彫帣偰丄BCG傪愙庬偡傞偲丄愙庬嬊強偑丄愙庬偟偰10擔埲撪乮4乣5擔埲撪乯偵丄愒偔壔擽偟偰棃傞乮僐僢儂尰徾乯丅寢妀嬠偵姶愼偟偰偄側偄恖偑巒傔偰BCG愙庬傪庴偗傞偲丄愙庬10擔屻崰偐傜丄恓嵀晹埵偵敪愒偑惗偠丄愙庬1乣2寧屻崰傑偱丄壔擽憙偑巆傞丅婛偵寢妀嬠偵姶愼偟偰偄傞恖乮彫帣乯偑偰BCG愙庬傪庴偗傞偲丄僐僢儂尰徾偵傛傝丄愙庬10擔屻埲撪偵丄愙庬嬊強偺敪愒丒庮挴傗丄恓嵀晹埵偺壔擽摍偑尰傟傞偑丄捠忢2乣4廡娫屻偵丄徚墛丄釐嵀壔偟丄帯桙偡傞偲尵傢傟傞丅
丂BCG愙庬傪巒傔偰庴偗偨彫帣偺応崌丄僣儀儖僋儕儞斀墳偑梲惈壔乮梲揮乯偡傞偵偼丄BCG愙庬屻6乣8廡娫屻偲尵傢傟傞丅
丂傕偟丄BCG愙庬傪庴偗偨巕嫙偑丄僐僢儂尰徾偲巚傢傟傞敪愒側偳偑尰傟偰偨応崌偵偼丄BCG愙庬2廡娫埲撪偵丄僣儀儖僋儕儞斀墳傪峴偆偲丄婛偵丄寢妀嬠偵姶愼偟偰偄偨偺偐丄妋擣弌棃傞偲尵偆丅
丂嶲峫暥專
丂丒嶳杮堦旻丄懠丗僇儔乕恾夝丂恱懷偺惓忢峔憿偲婡擻丂俬倁丂寣塼丒柶塽丒撪暘斿丂乮擔杮堛帠怴曬幮丄2002擭乯丏丂
丂丒墦揷墄巕丄懠丗僒僀僩僇僀儞偲働儌僇僀儞丂傢偐傝傗偡偄柶塽幘姵丂擔杮堛巘夛嶨帍丂摿暿崋乮1乯丂S46-S50, 2005擭丏
丂丒彫椦搊丗椪彴堛偺偨傔偺柶塽壢妛丂擔杮堛巘夛嶨帍丂惗奤嫵堢僔儕乕僘亅俆丂戞93姫丒戞8崋丄徍榓60擭丏
丂丒扟岥崕丄懠丗昗弨柶塽妛乮戞俀斉丄堛妛彂堾丄2004擭乯丏
丂丒桍戲岝旻丄懠丗攟梴僸僩T4+T嵶朎偺宍懺偲婡擻亅Recombinant IL-2偲傾儘峈尨巋寖偵傛傞挿婜攟梴亅丂慻怐攟梴丂14(3): 26-30, 1988擭丏
丂丒徏塝晀梇丗挿栰導堛巘夛抧堟梊杊愙庬峀堟壔悇恑偺偨傔偺尋廋夛丄挿栰缥曬丄暯惉18擭10寧1擔乮戞532崋乯丄22-32暸丏
丂丒梊杊愙庬僈僀僪儔僀儞丄梊杊愙庬儕僒乕僠僙儞僞乕丄2006擭3寧夵掶斉丏
丂http://idsc.nih.go.jp/vaccine/2006vagl/index.html