丂
丂 丂
丂 丂
丂
丂丂丂
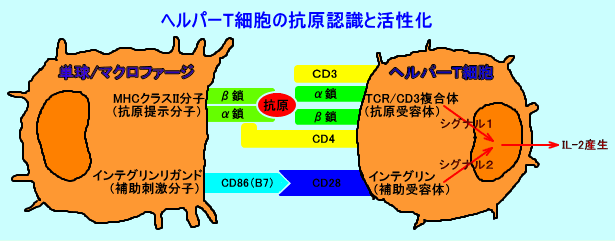
丂僿儖僷乕T嵶朎
丂T嵶朎昞柺偺TCR丒CD3暋崌懱偼丄扨媴丒儅僋儘僼傽乕僕昞柺偺丄峈尨偑寢崌偟偨MHC僋儔僗II暘巕傪丄擣幆偡傞丅
丂僿儖僷乕T嵶朎偺妶惈壔乮IL-2嶻惗乯偵偼丄峈尨採帵嵶朎偺MHC僋儔僗II暘巕亄峈尨儁僾僠僪偵傛傝丄僿儖僷乕T嵶朎偺TCR丒CD3暋崌懱偑丄峈尨摿堎揑偵巋寖偝傟傞偙偲乮僔僌僫儖侾乯偲丄峈尨採帵嵶朎偺曗彆巋寖暘巕乮僀儞僥僌儕儞儕僈儞僪乯偵傛傝丄僿儖僷乕T嵶朎偺曗彆庴梕懱乮僀儞僥僌儕儞乯偑丄峈尨旕摿堎揑偵巋寖偝傟傞偙偲乮僔僌僫儖俀乯偑丄昁梫丅
丂僿儖僷乕T嵶朎偼丄峈尨採帵嵶朎偵傛傝丄峈尨摿堎揑偵TCR丒CD3暋崌懱偑巋寖偝傟傞乮僔僌僫儖侾偱巋寖偝傟傞乯偩偗偱丄曗彆庴梕懱偑巋寖偝傟側偄乮僔僌僫儖俀偱巋寖偝傟側偄乯偲丄妶惈壔偝傟偢丄峈尨摿堎揑側柶塽姲梕偑丄桿摫偝傟傞丅
丂曗彆巋寖暘巕乮CD86乯偼丄峈尨採帵嵶朎乮儅僋儘僼傽乕僕側偳乯偺傒偑敪尰偟偰偄傞丅枛徑慻怐偺嵶朎偑丄MHC僋儔僗II暘巕偵丄帺屓峈尨傪寢崌偟偰丄帺屓峈尨傪採帵乮僔僌僫儖侾乯偟偰傕丄僿儖僷乕T嵶朎偼丄曗彆庴梕懱乮CD28乯偺巋寖乮僔僌僫儖俀乯偑側偄偺偱丄妶惈壔偝傟偢丄柍斀墳乮anergy乯偵廔傢傞丅偙偺傛偆側婡彉偱丄僿儖僷乕T嵶朎偼丄乽帺屓乿偺惉暘乮帺屓峈尨乯偵偼丄柶塽墳摎乮帺屓柶塽乯傪婲偙偝側偄丅
丂侾丏扨媴丒儅僋儘僼傽乕僕偼丄僿儖僷乕T嵶朎偵峈尨傪採帵偡傞
丂嵶嬠側偳偼丄扨媴丒儅僋儘僼傽乕僕偵傛傝婷怘偝傟丄儕僜僜乕儉偱丄峈尨乮峈尨儁僾僠僪乯偵暘夝偝傟傞丅
丂峈尨乮峈尨儁僾僠僪乯偼丄MHC僋儔僗II暘巕乮拲侾乯偺峚偵嫴傑傟傞傛偆偵寢崌偟偰丄扨媴丒儅僋儘僼傽乕僕昞柺偵採帵偝傟傞丅
丂扨媴丒儅僋儘僼傽乕僕昞柺偺MHC僋儔僗II暘巕偑丄峈尨乮峈尨儁僾僠僪乯傪嫴傓宍偱丄CD4梲惈乮拲俀乯偺僿儖僷乕T嵶朎偺TCR丒CD3暋崌懱偵峈尨傪採帵偡傞丅
丂峈尨採帵嵶朎乮峈尨掓帵嵶朎乯偱偁傞扨媴丒儅僋儘僼傽乕僕乮M兂乯偼丄嵶嬠傪婷怘偟丄嵶朎撪乮僄儞僪僝乕儉乯偱徚壔偟丄嵶嬠偺抈敀傪丄嵶偐偔嵱偒乮暘夝丒抐曅壔偟乯丄傾儈僲巁悢偑栺侾侽乣侾俆掱搙偺儁僾僠僪乮peptide乯偵偡傞丅庽忬嵶朎傗B嵶朎傕丄峈尨採帵偡傞丅
丂偙偆偟偰丄儁僾僠僪乮peptide乯偺宍偵偝傟偨峈尨儁僾僠僪乮峈尨僼儔僌儊儞僩丄儁僾僠僪抐曅丄峈尨抐曅丄儁僾僠僪峈尨乯偼丄扨媴丒儅僋儘僼傽乕僕偺MHC僋儔僗俬俬暘巕偵嫴傑傟傞傛偆偵偟偰丄僿儖僷乕T嵶朎偺TCR丒CD3暋崌懱偵丄採帵乮processing乯偝傟傞丅
丂TCR丒CD3暋崌懱偼丄TCR乮T嵶朎庴梕懱丗拲俁乯偑丄CD3暘巕偲嵶朎枌娧捠晹偱丄旕嫟桳揑偵寢崌偟偰偄傞丅彫朎懱偱崌惉偝傟偨TCR偺兛嵔丄兝嵔丄CD3暘巕偺億儕儁僾僠僪兞丄兟丄兠丄兡偼丄兛兝/兞兠/兟兠/兡-兡傪宍惉偟偰丄嵶朎枌偺TCR丒CD3暋崌懱偲偟偰丄敪尰偡傞丅
丂TCR偺峈尨擣幆偺僔僌僫儖偼丄CD3暘巕傪夘偟偰丄T嵶朎撪偵揱払偝傟傞乮僔僌僫儖侾乯丅
丂僿儖僷乕T嵶朎偑丄MHC僋儔僗II暘巕偵寢崌偟偨峈尨傪擣幆偡傞嵺偵偼丄CD4峈尨偑丄曗彆揑偵摥偔丅
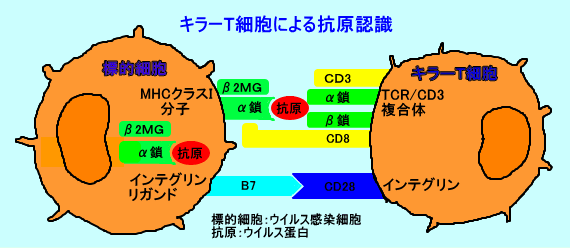
丂僂僀儖僗側偳偺峈尨偼丄峈尨儁僾僠僪乮峈尨僼儔僌儊儞僩乯偺宍偱丄扨媴丒儅僋儘僼傽乕僕偺MHC僋儔僗俬暘巕偲嫟偵丄CD8梲惈偺僉儔乕T嵶朎乮CTL丗cytotoxic T lymphocyte乯偵丄採帵偝傟傞丅
丂扨媴丒儅僋儘僼傽乕僕偺懠偵丄妶惈壔B嵶朎丄旂晢偺儔儞僎儖僴儞僗嵶朎丄慻怐撪偺庽忬嵶朎乮dendritic cell乯側偳傕丄峈尨採帵偡傞乮拲俆乯丅
丂T嵶朎偑妶惈壔偝傟傞偨傔偵偼丄TCR丒CD3暋崌懱傪夘偡傞峈尨摿堎揑側僔僌僫儖偵傛傞巋寖偲丄僀儞僥僌儕儞傪夘偡傞峈尨旕摿堎揑側僔僌僫儖偵傛傞嫟巋寖乮暃巋寖乯偑昁梫丅
丂侾乯丏僔僌僫儖侾丗MHC僋儔僗II暘巕乮峈尨採帵嵶朎乯/峈尨儁僾僠僪/TCR丒CD3暋崌懱乮T嵶朎乯偺峈尨擣幆僔僌僫儖
丂僿儖僷乕T嵶朎偼丄TCR丒CD3暋崌懱偑丄扨媴丒儅僋儘僼傽乕僕昞柺偺MHC僋儔僗俬俬暘巕傪夘偟偰丄峈尨儁僾僠僪傪採帵偝傟傞偙偲偱巋寖偝傟惗偠傞僔僌僫儖侾乮TCR僔僌僫儖丗峈尨採帵僔僌僫儖乯偩偗偱偼丄妶惈壔偝傟側偄丅
丂僔僌僫儖侾偼丄TCR丒CD3暋崌懱傪夘偡傞峈尨摿堎揑側僔僌僫儖丅
丂俀乯丏僔僌僫儖俀丗僀儞僥僌儕儞乮T嵶朎乯/僀儞僥僌儕儞儕僈儞僪乮峈尨採帵嵶朎乯偺暃僔僌僫儖
丂僿儖僷乕T嵶朎偼丄T嵶朎昞柺偺僀儞僥僌儕儞乮曗彆庴梕懱乯偑丄扨媴丒儅僋儘僼傽乕僕昞柺偵敪尰偝傟傞僀儞僥僌儕儞儕僈儞僪偱巋寖偝傟惗偠傞僔僌僫儖俀乮嫟巋寖僔僌僫儖丗曗彆巋寖僔僌僫儖乯傕丄T嵶朎妶惈壔偺偨傔偵昁梫丅
丂僔僌僫儖俀偼丄曗彆庴梕懱傪夘偡傞峈尨旕摿堎揑側僔僌僫儖丅
丂僿儖僷乕T嵶朎偺妶惈壔偵偼丄僔僌僫儖侾偲僔僌僫儖俀偺俀偮偺僔僌僫儖偑昁梫丅
丂僿儖僷乕T嵶朎偑丄峈尨採帵嵶朎乮峈尨掓帵嵶朎乯偺MHC僋儔僗II暘巕亄峈尨儁僾僠僪偵傛傝丄僿儖僷乕T嵶朎偺TCR丒CD3暋崌懱偑丄峈尨摿堎揑偵巋寖偝傟傞乮僔僌僫儖侾乯偲丄峈尨採帵嵶朎偺僀儞僥僌儕儞儕僈儞僪乮B7-1乯偵傛傝丄僿儖僷乕T嵶朎偺曗彆庴梕懱乮僀儞僥僌儕儞丗CD28傗CTLA-4乯偑丄峈尨旕摿堎揑偵巋寖偝傟傞乮僔僌僫儖俀乯偲丄僿儖僷乕T嵶朎偺嵶朎幙偱丄IL-2 mRNA偺揮幨偑婲偙傝丄IL-2偑惗嶻偝傟丄T嵶朎偼丄妶惈壔偝傟傞乮嫅愨斀墳偑婲偙傞乯丅
丂俁丏俀僔僌僫儖儌僨儖
丂戞侾偵丄T嵶朎偼丄峈尨擣幆偺僔僌僫儖侾偑側偔丄僔僌僫儖俀偑揱偊傜傟偰傕丄妶惈壔偝傟側偄丅
丂戞俀偵丄T嵶朎偼丄峈尨擣幆偺僔僌僫儖侾偺傒偑揱傢傝丄僔僌僫儖俀偑揱偊傜傟側偄偲丄妶惈壔偝傟側偄偩偗偱側偔丄偦偺峈尨偵懳偟偰丄墳摎偟側偄丄柶塽姲梕忬懺偵側傞丅
丂峈僀儞僥僌儕儞峈懱傪丄儅僂僗偺怱憻堏怉帪偵搳梌偡傞偲丄嫅愨斀墳偑梷惂偝傟傞丅屻偵丄偙偺儅僂僗偵丄摨偠僪僫乕儅僂僗偺旂晢傪堏怉偡傞偲丄峈僀儞僥僌儕儞峈懱傪搳梌偟側偔偰傕丄嫅愨斀墳側偟偵丄旂晢偼惗拝偟偨丅
丂儅僂僗偱偼丄堏怉捈屻偵丄CD4丄LFA-1偵懳偡傞峈懱傪丄抁婜娫搳梌偡傞偲丄堏怉偟偨慻怐偺傾儘峈尨偵摿堎揑側柶塽姲梕偑惉棫偟丄堏怉憻婍偑丄塱媣偵惗拝偡傞丅
丂係丏嫟巋寖暘巕乮僀儞僥僌儕儞乯偲丄懳墳偡傞儕僈儞僪
丂嫟巋寖暘巕乮暃巋寖暘巕丗曗彆庴梕懱乯偲偟偰丄CD28傗丄CD152乮CTLA-4乯丄CD40側偳偲尵偆丄僀儞僥僌儕儞乮庴梕懱乯偑丄T嵶朎昞柺偵敪尰偝傟傞丅
丂僀儞僥僌儕儞乮嫟巋寖暘巕丗曗彆庴梕懱乯偼丄懳墳偡傞儕僈儞僪乮曗彆巋寖暘巕乯偲丄嫟巋寖宱楬乮暃巋寖宱楬乯傪峔惉偡傞丅
丂僀儞僥僌儕儞乮曗彆庴梕懱乯/儕僈儞僪乮曗彆巋寖暘巕乯慻傒崌偣
丂侾乯丏CD28/B7嫟巋寖宱楬
丂T嵶朎昞柺偺CD28乮曗彆庴梕懱乯偑丄扨媴丒儅僋儘僼傽乕僕乮峈尨採帵嵶朎乯昞柺偵敪尰偝傟傞B7乮僀儞僥僌儕儞儕僈儞僪丗曗彆巋寖暘巕乯偲寢崌偡傞偲丄僔僌僫儖俀偑揱偊傜傟偰丄T嵶朎偼丄妶惈壔偝傟傞丅
丂B7曗彆巋寖暘巕乯偼丄摐抈敀偱丄B7-1乮CD80乯丄B7-2乮CD86乯偲尵偆丄俀偮偺暘巕偺堚揱巕偺懚嵼偑丄抦傜傟偰偄傞丗T嵶朎乮CD4梲惈僿儖僷乕T嵶朎乯偺僀儞僥僌儕儞/峈尨採帵嵶朎偺儕僈儞僪偺憡屳嶌梡偲偟偰丄CD28/CD80偲丄CD28/CD86偲偑偁傞丅
丂CD28偼丄CD3梲惈T嵶朎偺戝敿偑丄敪尰偡傞丅CD4梲惈T嵶朎偺曽偑丄CD8梲惈T嵶朎傛傝丄崅昿搙偵CD28峈尨傪敪尰偡傞丅
丂CD28偼丄妶惈壔傪桿摫偡傞惓偺僔僌僫儖傪揱払偡傞丅
丂CD86偺傛偆側僀儞僥僌儕儞儕僈儞僪偼丄僿儖僷乕T嵶朎偺曗彆庴梕懱傪巋寖偟丄僿儖僷乕T嵶朎傪妶惈壔偝偣傞丅
丂CD86偼丄峈尨採帵嵶朎乮B嵶朎丄庽忬嵶朎丄儅僋儘僼傽乕僕乯偺昞柺偵偟偐敪尰偟偰偄側偄丅枛徑慻怐偺嵶朎偑丄MHC僋儔僗II暘巕偵丄乽帺屓乿偺峈尨偺抐曅乮帺屓峈尨乯傪寢崌偟偰丄帺屓峈尨傪採帵偟偰傕丄僿儖僷乕T嵶朎偼丄僔僌僫儖俀乮CD28偵傛傞曗彆庴梕懱偺巋寖僔僌僫儖乯偑側偄偺偱丄妶惈壔偝傟偢丄柍斀墳乮anergy乯偵廔傢傞丅偙偺傛偆側婡彉偱丄僿儖僷乕T嵶朎偼丄乽帺屓乿偺惉暘乮帺屓峈尨乯偵偼丄柶塽墳摎乮帺屓柶塽乯傪婲偙偝側偄丅
丂俀乯丏CD152乮CTLA-4乯/B7嫟巋寖宱楬
丂T嵶朎昞柺偺CD152乮CTLA-4乯偑丄扨媴丒儅僋儘僼傽乕僕昞柺偺B7偲寢崌偡傞偲丄僿儖僷乕T嵶朎偺柶塽斀墳乮俬俴-2嶻惗側偳乯傪梷惂偡傞偲峫偊傜傟偰偄傞丅
丂T嵶朎乮CD4梲惈僿儖僷乕T嵶朎乯偺僀儞僥僌儕儞/峈尨採帵嵶朎偺儕僈儞僪偺憡屳嶌梡偲偟偰丄CD152乮CTLA-4乯/CD80偲丄CD152乮CTLA-4乯/CD86偲偑偁傞丅
丂CD152乮CTLA-4乯偼丄妶惈壔傪梷惂偡傞晧偺僔僌僫儖傪揱払偡傞丅
丂俁乯丏CD40/CD154乮CD40L乯嫟巋寖宱楬
丂CD40乮僀儞僥僌儕儞丗庴梕懱乯偼丄B嵶朎丄扨媴丒儅僋儘僼傽乕僕丄庽忬嵶朎丄寣娗撪旂嵶朎偵敪尰偟偰偄傞丅
丂CD40偼丄庮釃夡巰場巕庴梕懱僼傽儈儕乕乮TNF receptor family乯偵懏偡傞枌抈敀丅
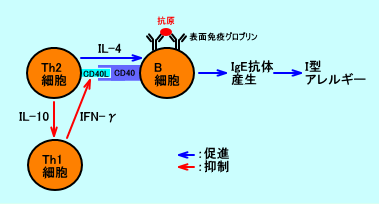
丂CD40儕僈儞僪乮CD4-ligand乯偺CD154乮CD40L丄gp39乯偼丄妶惈壔CD4梲惈T嵶朎昞柺乮僿儖僷乕T嵶朎偱偁傞Th2嵶朎乯偵敪尰偟偰偄傞丗峈尨採帵嵶朎偺僀儞僥僌儕儞/T嵶朎乮CD4梲惈僿儖僷乕T嵶朎乯偺儕僈儞僪偺憡屳嶌梡偲偟偰丄CD40/CD40L乮CD154乯偑偁傞丅
丂CD40偑丄CD154乮CD40L乯偲寢崌偡傞偲丄峈尨旕摿堎揑側僔僌僫儖乮僔僌僫儖俀乯偑揱払偝傟丄B嵶朎偺峈懱嶻惗偺僋儔僗僗僀僢僠乮拲係乯丄儕儞僷鄅朎偵堏摦偟偰偺泱拞怱偺宍惉丄婰壇B嵶朎偺弌尰偑桿摫偝傟傞丅
丂傑偨丄CD40/CD154乮CD40L乯嫟巋寖宱楬偼丄峈尨採帵嵶朎乮扨媴丒儅僋儘僼傽乕僕丄庽忬嵶朎乯傪妶惈壔偝偣丄峈尨採帵嵶朎昞柺偵嵶朎愙拝暘巕偺儕僈儞僪乮B7-1丄B7-2丄ICAM-1側偳乯傪敪尰偝偣偨傝丄峈尨採帵嵶朎偵墛徢惈僒僀僩僇僀儞乮拲俇乯側偳傪暘斿偝偣丄T嵶朎嫟巋寖宱楬乮CD28/B7嫟巋寖宱楬乯傪桿摫偡傞丅
丂偙偺傛偆偵丄CD40/CD154乮CD40L乯嫟巋寖宱楬偼丄B嵶朎偐傜偺峈懱嶻惗偺傒側傜偢丄T嵶朎偺妶惈壔偵傕丄廳梫側宱楬丅
丂堏怋柶塽偱丄CD154乮CD40L乯偵懳偡傞峈懱傪搳梌偡傞偲丄柶塽晄墳摎惈傪挿偔帩懕偝偣傞偙偲偑丄擣傔傜傟偰偄傞丅
丂IFN-兞偼丄CD40儕僈儞僪乮CD40L乯偺敪尰傪梷惂偟偰丄B嵶朎偵傛傞IgE峈懱嶻惗傪梷惂偡傞丅丂俆丏柶塽僔僫僾僗
丂T嵶朎偲峈尨採帵嵶朎偼丄捈愙丄愙拝偟丄憡屳嶌梡偟偰丄T嵶朎偑丄妶惈壔偝傟傞丅
丂愙拝偵嵺偟偰丄T嵶朎偲峈尨採帵嵶朎偺娫偵偼丄條乆側暘巕孮偑娭梌偟丄柶塽僔僫僾僗偑丄宍惉偝傟傞丅
丂曗彆庴梕懱/儕僈儞僪偲偟偰偼丄LFA-1/ICAM-1丄CD28/CD80乮B7-1乯丄CD152乮CTLA-4乯/CD80乮B7-1乯側偳偑丄宍惉偝傟丄摨庬堏怉柶塽偺媫惈嫅愨斀墳偵娭梌偡傞丅
丂昞侾丂T嵶朎偲峈尨採帵嵶朎偺憡屳嶌梡 丂T嵶朎丂 丂峈尨採帵嵶朎丂 丂TCR丒CD3丒CD4乮CD8乯 丂MHC亄峈尨儁僾僠僪 丂CD2乮LFA-2乯 丂CD58乮儅僂僗偱偼CD48乯 丂CD28 丂CD80乮B7-1乯丄CD86乮B7-2乯 丂CD152乮CTLA-4乯 丂CD80乮B7-1乯丄CD86乮B7-2乯 丂CD11a/CD18乮LFA-1乯 丂CD54乮ICAM-1乯 丂CD40L乮CD154乯 丂CD40 丂CD43丄CD45 丂- 丂CD2乮LFA-2乯偼丄T嵶朎傗NK嵶朎偑丄梤偺愒寣媴乮SRBC乯偲丄E-儘僛僢僩傪宍惉偡傞嵺偺丄儕僙僾僞乕丅側偍丄T嵶朎昞柺偺CD2乮僀儞僥僌儕儞乯偑寢崌偡傞丄峈尨採帵嵶朎昞柺偺儕僈儞僪偼丄僸僩偱偼丄CD58乮LFA-3乯偱丄儅僂僗偱偼丄CD48丗儅僂僗偱偼丄CD2偼丄CD48偺傒傪丄儕僈儞僪偲偡傞丅僸僩偱偼丄CD2偼丄CD48偵傕丄庛偄恊榓惈傪帵偡偑丄CD58乮LFA-3乯偵丄崅偄恊榓惈傪帵偡丅
丂CD3偼丄TCR偲丄TCR丒CD3暋崌懱傪宍惉偡傞丅
丂CD4傗CD8偼丄TCR偑丄MHC亄峈尨儁僾僠僪傪擣幆偡傞嵺丄MHC暘巕偵寢崌偟丄TCR偺MHC亄峈尨儁僾僠僪偲偺寢崌傪丄埨掕壔偝偣傞丅
丂CD11a/CD18乮LFA-1丗lymphocyte function-associated antigen乯偼丄僀儞僥僌儕儞偲偟偰丄儕僈儞僪偱偁傞ICAM-1乮intercellular adhesion molecule-1乯偲丄寢崌偡傞丅CD11a/CD18乮LFA-1乯偺儕僈儞僪偲偟偰偼丄ICAM-1埲奜偵丄ICAM-2乮CD102乯丄ICAM-3乮CD50乯偑丄懚嵼偡傞丅LFA-1偼丄敀寣媴傗儕儞僷慻怐偵偩偗敪尰偟丄ICAM-1偼丄B嵶朎傗峈尨採帵嵶朎乮APC丗扨媴傗庽忬嵶朎乯側偳偺寣塼宯嵶朎偲丄寣娗撪旂嵶朎偵敪尰偡傞丅LFA-1乮僀儞僥僌儕儞乯偲ICAM-1乮儕僈儞僪乯偲偺寢崌偵傛傝丄庬乆偺嵶朎偑丄愙拝偡傞丅
丂CD28偼丄T嵶朎傪丄惓偵惂屼偡傞丗T嵶朎偼丄CD28偵傛傝丄峈尨採帵嵶朎偐傜偺僔僌僫儖俀乮戞擇僔僌僫儖丗曗彆庴梕懱僔僌僫儖乯傪庴梕偟丄妶惈壔偝傟丄IL-2傗IL-2R乮IL-2庴梕懱乯傪敪尰偟丄憹怋偡傞丅T嵶朎偼丄TCR丒CD3暋崌懱傪夘偡傞僔僌僫儖侾偵傛傝巋寖偝傟偰傕丄CD28暘巕偑丄峈尨採帵嵶朎偺曗彆庴梕懱乮CD80傗CD86乯偐傜偺僔僌僫儖俀偵傛傝巋寖偝傟側偄偲丄妶惈壔偝傟偢乮IL-2mRNA偺敪尰偑憹壛偟側偄乯丄柶塽姲梕偑桿摫偝傟傞丅IL-2mRNA偺敪尰偼丄T嵶朎偑丄TCR丒CD3暋崌懱傪夘偡傞僔僌僫儖侾偵傛傝巋寖偝傟傞偲丄庒姳検憹壛偡傞偑丄峈尨採帵嵶朎偺曗彆庴梕懱偐傜偺僔僌僫儖俀偵傛傝巋寖偝傟傞偲丄5乣20攞偵憹壛偟丄IL-2嶻惗検偑憹壛偡傞丅
丂CD152乮CTLA-4乯偼丄T嵶朎傪丄晧偵惂屼偡傞丗CD152乮CTLA-4乯偼丄CD28偵傛傞T嵶朎傊偺惓偺巋寖傪丄挷愡偟偰偄傞偲丄峫偊傜傟偰偄傞丅
丂CD152乮CTLA-4乯偺峔憿偼丄CD28偵椶帡偟丄CD152乮CTLA-4乯偺儕僈儞僪傕丄CD28偲摨偠丄CD80乮B7-1乯傗CD86乮B7-2乯丅
丂CD152乮CTLA-4乯偼丄媥巭婜乮惷巭婜乯T嵶朎偵敪尰偣偢丄妶惈壔偝傟偨T嵶朎偑丄敪尰偡傞丅
丂CD152乮CTLA-4乯偼丄CD4梲惈T嵶朎乮僿儖僷乕T嵶朎乯偑丄帺屓斀墳惈T嵶朎傪梷惂偡傞嵺偵昁梫偱丄帺屓柶塽幘姵偺敪徢偺梷惂偵娭梌偟偰偄傞偲丄峫偊傜傟偰偄傞丅
丂CD80乮B7-1乯偼丄妶惈壔B嵶朎傗丄妶惈壔儅僋儘僼傽乕僕偑丄敪尰偡傞丅
丂CD80乮B7-1乯偼丄CD28傗CD152乮CTLA-4乯偺儕僈儞僪偱偁傞丅CD80乮B7-1乯埲奜偵傕丄CD86乮B7-2乯傕丄CD28傗CD152乮CTLA-4乯偺儕僈儞僪偱偁傞丅CD80乮B7-1乯傗CD86乮B7-2乯偼丄CD28傛傝傕丄CD152乮CTLA-4乯偵丄傛傝嫮偔寢崌偡傞乮恊榓椡偑嫮偄乯丅
丂CD80乮B7-1乯傗CD86乮B7-2乯偼丄峈尨採帵嵶朎偑丄敪尰偡傞丗CD80乮B7-1乯偼丄庽忬嵶朎偑丄敪尰偡傞丅CD80乮B7-1乯偼丄媥巭婜B嵶朎偼丄敪尰偣偢丄妶惈壔B嵶朎偑丄敪尰偡傞丅CD80乮B7-1乯偼丄妶惈壔儅僋儘僼傽乕僕偑丄敪尰偡傞丅CD86乮B7-2乯偼丄媥巭婜B嵶朎傕丄敪尰偡傞丅丂丂
丂CD45偼丄LCA乮Leukocyte common antigen乯丄T200偲柤偯偗傜傟偨昞柺峈尨偱丄摐嵔偵晉傫偱偄傞丅
丂CD45乮LCA乯偵偼丄CD45RA丄CD45RO偲尵偆丄isoform偑懚嵼偡傞丅CD4梲惈嵶朎偱丄CD45RA傪敪尰偟偰偄傞嵶朎乮CD45RA梲惈嵶朎乯偼丄僒僾儗僢僒乕/僀儞僨儏乕僒乕婡擻乮S/I婡擻乯傪桳偟丄CD45RO傪敪尰偟偰偄傞嵶朎偼丄僿儖僷乕/僀儞僨儏乕僒乕婡擻乮H/I婡擻乯傪桳偟偰偄傞丅
丂CD45偼丄NK嵶朎偵傕丄敪尰偝傟偰偄傞丅CD45偵懚嵼偡傞摐嵔乮poly-N-acetyllactosamine丗LacNAc乯偼丄NK嵶朎偺昗揑嵶朎偲偺寢崌傪丄梷惂乮慾奞乯偡傞丅
丂俇丏Hygiene hypothesis
丂儕億懡摐乮LPS丗lipopolysaccharide丄僌儔儉堿惈嬠乯丄CpG僆儕僑僰僋儗僆僠僪丄儕億儁僾僠僪丄儁僾僠僪僌儕僇儞乮peptideglycan丗僌儔儉梲惈嬠乯丄儕億傾儔價僲儅儞僫儞乮lipoarabinomannan丄寢妀嬠乯側偳偺旝惗暔偺嬠懱惉暘偼丄庽忬嵶朎偺Toll-like receptor 乮拲俈乯傪夘偟偰丄庽忬嵶朎傪妶惈壔偝偣丄CD40丄CD80丄CD86側偳偺嫟巋寖暘巕乮暃巋寖暘巕乯傪敪尰偝偣傞乮NF-kB偑妶惈壔偝傟傞乯丅僫僀乕僽T嵶朎偼丄庽忬嵶朎偐傜採帵偝傟傞峈尨丄嫟巋寖暘巕乮暃巋寖暘巕乯丄墛徢惈僒僀僩僇僀儞乮IL-12丄TNF-兛乯偵傛傝妶惈壔偝傟傞偲丄Th1嵶朎偑丄桪埵偵暘壔偡傞丅Th1嵶朎偼丄IFN-兞傪嶻惗偟丄IgE峈懱偺嶻惗傪梷惂偟丄傾儗儖僊乕斀墳傪梷惂偡傞偲尵傢傟傞丅
丂俈丏偦偺懠偺愙拝暘巕丂VLA-4/VCAM-1偺憡屳嶌梡傕丄僿儖僷乕T嵶朎偺妶惈壔偵廳梫偱丄儅僂僗偱偺怱憻堏怉偺幚尡偱丄VCAM-1傗VLA-4偵懳偡傞峈懱傪搳梌偡傞丄堏怋偟偨怱憻偵嫅愨斀墳偑婲偙傜側偄偲尵偆丅
丂俉丏帺屓峈懱嶻惗偺epitope spreading壖愢
丂帺屓柶塽幘姵偱偼丄帺屓峈尨偲斀墳偡傞丄帺屓峈懱偑嶻惗偝傟偰偄傞丅
丂懡偔偺帺屓峈尨偺傾儈僲巁攝楍偼丄僂僀儖僗傗嵶嬠抈敀偺堦晹偲丄暘巕儗儀儖偱憡摨偟偰偄傞偙偲偑丄妋擣偝傟偨丅
丂椺偊偽丄U1RNP-70K抈敀偲尵偆帺屓峈尨偼丄B宆僀儞僼儖僄儞僓僂僀儖僗偺僂僀儖僗峈尨偲丄憡摨惈偑偁傞丅
丂epitope spreading壖愢偵傛傞偲丄傑偢丄僂僀儖僗姶愼偱嶻惗偝傟偨丄帺屓峈尨偵椶帡偟偨僂僀儖僗峈尨偵懳偡傞峈懱偼丄岎嵎斀墳偱丄帺屓峈尨乮A僄僺僩乕僾乯偲傕斀墳偡傞丅
丂偙偺丄帺屓峈尨偵椶帡偟偨僂僀儖僗峈尨偵懳偡傞峈懱傪儕僙僾僞乕偵寢崌偟偨B嵶朎偼丄帺屓峈尨傪T嵶朎偵峈尨採帵偡傞丅
丂偦偆偡傞偲丄帺屓峈尨乮A僄僺僩乕僾乯嬤朤偺丄怴偨側帺屓峈尨乮B僄僺僩乕僾乯偵懳墳偡傞B嵶朎僋儘乕儞傕妶惈壔偝傟丄恀偺帺屓峈懱偑嶻惗偝傟傞偲峫偊傜傟傞丅
丂epitope spreading壖愢偼丄儅僂僗偺峈U1RNP峈懱嶻惗宯偱妋擣偝傟偨偲偄偆丅
丂偙偺傛偆偵丄帺屓峈尨偲憡摨惈傪帩偮昦尨旝惗暔偺姶愼傪宊婡偵丄帺屓峈懱偑嶻惗偝傟傞偲傕峫偊傜傟傞丅
丂惗懱偼丄旝惗暔偵姶愼偡傞偲丄旝惗暔偑桳偡傞抈敀乮抈敀傗懡摐懱亖峈尨乯偵懳偟偰丄柶塽墳摎偟丄旝惗暔桼棃偺峈尨偵懳偟偰丄峈懱嶻惗側偳傪偍偙側偆丅摨偠丄旝惗暔偵懳偡傞柶塽墳摎乮峈懱嶻惗側偳乯偼丄廻庡乮僸僩乯偑桳偡傞MHC乮MHC僋儔僗II暘巕側偳乯偺宆偵傛偭偰丄堎側傞丅
丂廻庡乮僸僩乯偼丄帺屓偺慻怐偵丄旝惗暔偑桳偡傞抈敀乮抈敀傗懡摐懱亖峈尨乯偲帡偨峔憿偺抈敀側偳傪丄桳偟偰偄傞丅
丂姶愼偟偨旝惗暔偵懳偟偰丄崅墳摎偟偰丄嫮偄柶塽墳摎偑婲偙傞偲丄嶻惗偝傟偨旝惗暔桼棃偺峈尨偵懳偡傞峈懱側偳偑丄旝惗暔偑桳偡傞峈尨偲帡偨峔憿傪偟偨丄帺屓偺慻怐偺抈敀傪丄柶塽揑偵忈奞偡傞偙偲偑丄帺屓柶塽幘姵敪徢偺梫場偲峫偊傜傟偰偄傞丅
丂俋丏庽忬嵶朎
丂庽忬嵶朎乮dendritic cells乯偼丄峈尨採帵嵶朎偲偟偰丄廳梫丅
丂庽忬嵶朎偼丄MHC僋儔僗I暘巕傗丄MHC僋儔僗II暘巕傗丄曗彆庴梕懱乮僀儞僥僌儕儞乯偺儕僈儞僪乮CD86乯傪丄忢帪丄懡検偵丄敪尰偟偰偄傞丅
丂庽忬嵶朎帺恎丄僂僀儖僗偵姶愼偟堈偄丅
丂庽忬嵶朎偼丄崪悜偱暘壔偟偨屻丄枛徑慻怐偵寣塼拞傪堏峴偟丄婥摴丄徚壔娗擲枌丄旂晢偺昞旂偵丄暘晍偡傞丅昞旂偺庽忬嵶朎偼丄儔儞僎儖僴儞僗嵶朎乮Langerhans cells乯偲丄屇偽傟傞丅
丂庽忬嵶朎偼丄怘嶌梡偱丄峈尨傪庢傝崬傓偲丄嬤偔偺儕儞僷愡偵堏摦偟丄嵶朎幙撍婲傪怢挘偟丄楢寢偟丄interdiditating cell偲側傝丄峈尨採帵傪峴偆丅
丂庽忬嵶朎偼丄昦尨懱偺旝惗暔偑丄惗懱偵怤擖偡傞偲丄旝惗暔峈尨傪婷怘偡傞丅
丂庽忬嵶朎偼丄旝惗暔偑嶻惗偡傞暘巕乮儕億懡摐側偳乯傗丄嶻惗偝傟傞墛徢惈僒僀僩僇僀儞乮IL-1丄TNF-兛乯偵傛傝丄惉弉偟丄昞柺偵MHC亄峈尨儁僾僠僪暋崌懱偲丄曗彆巋寖暘巕乮CD80丄CD86乯傪丄崅儗儀儖偵敪尰偡傞丅
丂峈尨採帵嵶朎偱傕丄儅僋儘僼傽乕僕偼丄峈尨偲側傞昦尨懱側偳偑怤擖偡傞偲丄妶惈壔偝傟丄昦尨懱側偳傪婷怘偟丄怘朎撪偱暘夝偟丄MHC暘巕亄峈尨儁僾僠僪暋崌懱偲偟偰丄嵶朎昞柺偵丄敪尰偡傞丅
丂侾侽丏挵撪嵶嬠偲柶塽姲梕
丂挵撪嵶嬠乮忢嵼嵶嬠憄乯偼丄徚壔娗偺柶塽姲梕偵丄昁梫丅
丂儅僂僗偵懡検乮20乣25mg乯偺棏敀傾儖僽儈儞傪怘傋偝偣傞偲丄棏敀傾儖僽儈儞偵懳偟偰丄柶塽姲梕偵側傞丅柍嬠儅僂僗偱偼丄柶塽姲梕偑惉棫偟側偄偑丄5廡楊傑偱偵丄柍嬠儅僂僗偺徚壔娗撪偵丄忢嵼嵶嬠憄傪宍惉偝偣傞偲丄柶塽姲梕偑惉棫偡傞傛偆偵側傞丅
丂寬峃恖桼棃偺儕儞僷媴乮寬峃恖偺枛徑寣傗丄挵娗擲枌偐傜暘棧偝傟偨儕儞僷媴乯偼丄摨偠恖偺挵撪嵶嬠偺嬠懱傗丄拪弌暔偵傛偭偰偼丄巋寖偝傟側偄乮blast formation偑婲偙傜側偄乯丅偟偐偟丄僋儘乕儞昦乮Crohn's disease乯傗丄捵釃惈戝挵墛偺姵幰桼棃偺儕儞僷媴偼丄摨偠恖乮姵幰乯偺挵撪嵶嬠偺嬠懱傗丄嵶嬠偺昞柺峈尨偵傛傝丄巋寖偝傟傞丅偙偺傛偆偵丄僋儘乕儞昦傗丄捵釃惈戝挵墛偺姵幰偼丄徚壔娗偺柶塽姲梕偑惉棫偟偰偍傜偢丄挵娗撪偱丄挵撪嵶嬠乮忢嵼嵶嬠憄乯偵懳偟偰傕丄柶塽墳摎傪偟偰偟傑偆丅
丂僋儘乕儞昦傗捵釃惈戝挵墛偼丄媿擕傗擕惢昳偺愛庢傪嬛巭偟丄帺慠偺崚暔乮惛惢搙偑彮側偄崚暔乯丄栰嵷丄壥暔傪愛庢偝偣傞偲丄夵慞偟偨傝丄帯桙偡傞偲尵偆丅
丂徚壔娗偱偼丄塰梴慺傪媧廂偡傞嵺偵丄擲枌偼丄怘帠桼棃偺峈尨傗丄挵撪嵶嬠偺峈尨側偳丄庬乆偺峈尨偵敇偝傟偰偄傞丅徚壔娗偱偼丄嬊強柶塽偲偟偰丄IgA峈懱傪暘斿偟丄堎暔傪攔彍偟丄傑偨丄慡恎柶塽偲偟偰丄徚壔娗撪偺峈尨偵斀墳偟側偄傛偆偵丄柶塽姲梕傪桿摫偡傞丅偙偺傛偆側徚壔娗偺柶塽偼丄GALT乮gut-associated lymphoid tissue乯偲屇偽傟傞挵偺儕儞僷憰抲偱丄塩傑傟傞丅GALT偼丄僷僀僄儖斅乮Peyer斅乯丄挵娫枌儕儞僷愡丄挵擲枌屌桳憌傗忋旂嵶朎娫偺儕儞僷媴丄宍幙嵶朎丄旍枮嵶朎乮mast cell乯丄儅僋儘僼傽乕僕側偳偵傛傝峔惉偝傟傞丅GALT偼丄挵擲枌偺梕愊偺25亾傪愯傔傞丅
丂GALT偺僷僀僄儖斅偱偼丄僒僾儗僢僒乕T嵶朎乮Suppresor T cells乯傪桿摫偟偰丄怘塧惈抈敀峈尨傗丄挵撪嵶嬠偺嬠懱峈尨側偳偵懳偟偰丄柶塽姲梕乮宱岥姲梕乯傪惉棫偝偣傞偲尵傢傟傞偑丄偦偺婡彉偺徻嵶偼丄暋嶨偱丄夝柧偝傟偰偄側偄丅
丂挵撪嵶嬠乮忢嵼嵶嬠憄乯偺傛偆偵丄徚壔娗偺擲枌乮僶儕傾乕乯傪怤廝偟偰丄擲枌壓乮惗懱撪乯偵擖傝崬傑側偄峈尨偵懳偟偰偼丄柶塽姲梕乮宱岥姲梕乯偑桿摫偝傟傞丅偟偐偟丄昦尨惈嵶嬠側偳偵傛傝丄徚壔娗偺擲枌乮僶儕傾乕乯偑怤廝偝傟偨傝丄擲枌偺摟夁惈偑槾恑偟偰丄擲枌壓乮惗懱撪乯偵擖傝崬傫偩峈尨偵懳偟偰偼丄柶塽姲梕偑桿摫偝傟偢乮宱岥姲梕偑攋抅偟偰乯丄怘暔傾儗儖僊乕傗丄墛徢惈挵幘姵側偳偑丄婲偙傞丅
丂怘暔傾儗儖僊乕偼丄傾儗儖僊乕傪婲偙偡峈尨乮儁僾僠僪峈尨乯傪娷傓怘塧傪愛庢偡傞偙偲傛傝丄怘塧桼棃偺峈尨偑丄壗傜偐偺婡彉偱丄徚壔娗偺擲枌乮僶儕傾乕乯傪捠夁偟偰丄擲枌壓乮惗懱撪乯偵擖傝崬傓偙偲偑尨場偱丄婲偙傞丅廬偭偰丄傾儗儖僊乕傪婲偙偡峈尨偺彍嫀怘椕朄偼丄崻杮揑側帯椕偱偼側偄丅乮傓偟傠丄怘昳偵娷傑傟偰偄傞奅柺妶惈嵻側偳偺愛庢偺惂尷偺曽偑丄廳梫偲巚傢傟傞丅乯
丂挵娗偐傜偺帀朾偺媧廂偵敽側偄丄儕儞僷娗撪傪揮憲偝傟傞儕儞僷媴悢傕丄憹壛偡傞丅
丂惗懱尠旝嬀偱娤嶡偡傞偲丄挵娗峯撪偵丄僆儕乕僽桘傪搳梌偡傞偲丄挵娗偐傜偺帀朾媧廂乮帀朾偺揮憲乯偵敽側偭偰丄儕儞僷娗撪偵丄儕儞僷媴偑憹壛偡傞乮儕儞僷媴偺揮憲偑憹壛偡傞乯丅偙偺挵娗儕儞僷娗撪乮挵娗儕儞僷塼拞乯偺儕儞僷媴偼丄75亾偑丄CD4梲惈僿儖僷乕T嵶朎乮helper/inducer T cells乯丅
丂帀朾巁偱傕丄僆儗僀儞巁乮挿嵔晄朞榓帀朾巁乯丄僷儖儈僠儞巁乮挿嵔朞榓帀朾巁乯偼丄挵娗偐傜媧廂偝傟偨屻丄儕儞僷娗撪傪桝憲偝傟丄傑偨丄僇僾儕儖巁乮拞嵔帀朾巁乯偼丄栧柆撪傪桝憲偝傟傞丅僆儗僀儞巁乮挿嵔晄朞榓帀朾巁乯丄僷儖儈僠儞巁乮挿嵔朞榓帀朾巁乯偼丄乮怘帠偱愛庢偡傞偲丄乯挵娗儕儞僷娗撪偺儕儞僷媴悢傪丄憹壛偝偣傞乮儕儞僷媴偺揮憲傪丄憹壛偝偣傞乯丅偟偐偟丄僇僾儕儖巁乮拞嵔帀朾巁乯偼丄挵娗儕儞僷娗撪偺儕儞僷媴悢傪丄憹壛偝偣側偄丅僆儗僀儞巁傗僷儖儈僠儞巁側偳偺挿嵔帀朾巁偼丄儕儞僷媴梒庛壔斀墳乮blast formation乯傪丄槾恑偝偣傞乮挿嵔帀朾巁偼丄儕儞僷媴偺憹怋傗妶惈壔傪懀恑偝偣傞乯丅
丂惗懱偼丄僶儕傾乕乮僶儕傾乯偲偟偰偺挵娗忋旂傗旂晢傪墇偊偰怤擖偡傞昦尨懱偵偼丄柶塽墳摎傪偡傞丅
丂Toll條庴梕懱偺TLR5偼丄挵娗偺忋旂嵶朎偺栄嵶寣娗懁偵懚嵼偡傞丅惗懱偼丄挵娗偺忋旂嵶朎昞柺偵懚嵼偡傞挵撪嵶嬠乮忢嵼嵶嬠乯偵懳偟偰偼丄柶塽姲梕傪桿摫偡傞偑丄挵娗擲枌傪墇偊偰怤擖偟偨挵撪嵶嬠偵懳偟偰偼丄TLR5側偳偱丄擣幆偟乮姶抦偟乯丄柶塽墳摎傪偡傞丅
丂偍傑偗
丂儕儞僷媴傗岲拞媴乮梓棻媴乯偼丄儅僋儘僼傽乕僕乮扨媴丄僌儕傾嵶朎丄僋僢僷乕嵶朎側偳乯偐傜暘壔偟偨偲尵傢傟傞丅NK嵶朎傗丄寣娗撪旂嵶朎傕丄儅僋儘僼傽乕僕偐傜暘壔偟偨偺偱丄堎暔乮嵶嬠側偳乯傪婷怘偡傞惈幙偑偁傞丅
丂儕儞僷媴偼丄T嵶朎乮T儕儞僷媴乯偲丄B嵶朎乮B儕儞僷媴乯偲偵丄暘椶偝傟傞丅
丂T嵶朎乮T儕儞僷媴乯偼丄僿儖僷乕T嵶朎乮CD4梲惈丗峈尨採帵嵶朎偐傜峈尨傪採帵偝傟傞乯偲丄僉儔乕T嵶朎乮CD8梲惈丗僂僀儖僗姶愼嵶朎側偳傪摿堎揑偵嶦偡乯偲偵丄暘椶偝傟傞丅僿儖僷乕T嵶朎偼丄偝傜偵丄嵶朎惈柶塽偵娭梌偡傞Th1嵶朎偲丄塼惈柶塽偵娭梌偡傞Th2嵶朎偲偵暘椶偝傟傞丅
丂B嵶朎乮B儕儞僷媴乯偼丄僿儖僷乕T嵶朎偑暘斿偡傞僒僀僩僇僀儞偵傛傝丄宍幙嵶朎乮僾儔僘儅嵶朎乯偵暘壔偟丄峈懱乮柶塽僌儘僽儕儞乯傪嶻惗偡傞丅
丂屆偄柶塽宯偱偁傞嫻態奜暘壔T嵶朎傗丄B-1嵶朎偼丄帺屓墳摎惈柶塽嵶朎偱偁傝丄銹尨昦側偳偺帺屓柶塽幘姵偺敪徢偵娭梌偡傞丅
丂拲侾丗MHC偲偼丄major histocompatibility complex偺棯偱丄庡梫慻怐揔崌峈尨暋崌懱偺偙偲丅
丂恖娫偱偼丄HLA乮human leukocyte antigen乯偑丄MHC丅
丂堏怋偟偨慻怐偵丄嫅愨斀墳偑婲偙傞帪丄MHC偼丄乽旕帺屓乿偲偟偰擣幆偝傟偰偄傞丅
丂MHC乮HLA乯偺宆偵傛傝丄摿掕偺峈尨偵懳偡傞丄柶塽墳摎偺嫮搙偑堎側傞丅偙傟偼丄摨偠峈尨儁僾僠僪偱偁偭偰傕丄MHC乮HLA乯偺宆偵傛傝丄MHC乮HLA乯偺峚傊寢崌偟堈偐偭偨傝丄寢崌偟偵偔偄偨傔偲偝傟傞丅椺偊偽丄僸僩偺HLA偺DR51-B5偼丄僀儞僼儖僄儞僓僂僀儖僗偺HA偲丄嫮偔寢崌偡傞丅僸僩偺HLA偺DQ6-B5偼丄梟寣惈楢嵔媴嬠乮梟楢嬠乯偺M抈敀偲丄嫮偔寢崌偡傞丅廬偭偰丄HLA偺DR51-B5傪桳偡傞僸僩偼丄僀儞僼儖僄儞僓僂僀儖僗偵懳偟偰丄傑偨丄HLA偺DQ6-B5傪桳偡傞僸僩偼丄梟楢嬠偵懳偟偰丄嫮偄柶塽墳摎傪帵偡丅偙偺傛偆偵丄MHC乮HLA乯偺宆偵傛傝丄摿掕乮栤戣乯偺峈尨偲寢崌偺偟堈偝偵嵎偑偁傞丅MHC乮HLA乯偺宆偵傛傝丄摿掕偺峈尨偵懳偟偰丄柶塽墳摎偡傞偐丄柶塽旕墳摎側偺偐丄寛傑傞丅柶塽墳摎偑嫮偡偓傞乮崅偡偓傞乯偙偲偼丄壴暡徢側偳丄傾儗儖僊乕幘姵側偳傪棃偨偟丄惗懱偵晄棙側応崌傕偁傞丅
丂柶塽宯偱傕丄T嵶朎偼丄帺屓偺MHC乮HLA乯偑曄壔乮曄惈乯偟偰偄側偄偐丄擣幆偡傞丅T嵶朎偼丄TCR乮T嵶朎庴梕懱乯偵傛傝丄峈尨採帵嵶朎昞柺偺丄宆偺堎側傞MHC乮HLA乯傗丄曄惈偟偨MHC乮HLA乯傪丄乽旕帺屓乿偲偟偰擣幆偡傞偲峫偊傜傟傞丅
丂TCR乮T嵶朎庴梕懱乯偼丄MHC乮HLA乯偲峈尨儁僾僠僪偺暋崌懱乮MHC偲丄偦偺峚偵寢崌偟偨峈尨儁僾僠僪乯傪擣幆偡傞丅
丂MHC偼丄俀庬椶懚嵼偡傞丗
丂丒MHC僋儔僗俬暘巕丗兛嵔偲丄兝2儈僋儘僌儘僽儕儞偺兝嵔偱峔惉偝傟傞丅僸僩偱偼丄HLA-A丄B丄C峈尨偑懚嵼偡傞丅T嵶朎丄B嵶朎偺懠丄傎偲傫偳慡偰偺嵶朎昞柺傗丄寣彫斅昞柺偵丄敪尰偟偰偄傞丅嵶朎偵姶愼偟偨僂僀儖僗偼丄嵶朎幙撪偱僂僀儖僗抈敀幙傪崌惉偡傞丅僂僀儖僗抈敀幙偼丄僾儘僥傾僜乕儉偱丄峈尨儁僾僠僪乮傾儈僲巁偺悢偑俈乣俋働乯偵暘夝偝傟丄慹柺彫朎懱乮ER乯偵塣偽傟偰丄崌惉搑忋偺MHC僋儔僗俬暘巕偲寢崌偟偰丄嵶朎昞柺偵敪尰偝傟傞丅偙偺傛偆偵偟偰丄MHC僋儔僗俬暘巕偼丄嵶朎撪偱崌惉偝傟偨峈尨儁僾僠僪傪丄CD8梲惈僉儔乕T嵶朎偵採帵偡傞丅
丂僿儖儁僗僂僀儖僗側偳偺僂僀儖僗偼丄姶愼嵶朎偺MHC乮MHC僋儔僗I暘巕乯偺敪尰傪梷惂偟丄僉儔乕T嵶朎偺峌寕乮嵶朎忈奞妶惈乯傪摝傟傞丅偟偐偟丄MHC乮MHC僋儔僗I暘巕乯傪敪尰偟偰偄側偄嵶朎偼丄NK嵶朎偺峌寕傪庴偗傞丅
丂拲俀丗CD偲偼丄cluster of differentiation偺棯丅T嵶朎側偳偺柶塽婡峔傪宍惉偡傞嵶朎偺昞柺偵懚嵼偡傞丄婡擻揑偵廳梫側暘巕傪暘椶偡傞偺偵梡偄傜傟傞丅CD峈尨暘巕偼丄儌僲僋儘乕僫儖峈懱偱専嵏偝傟傞丅
丂拲俁丗TCR偲偼丄T cell receptor偺棯偱丄T嵶朎庴梕懱乮T嵶朎峈尨庴梕懱丄T嵶朎峈尨儗僙僾僞乕乯偺偙偲丅戝懡悢偺T嵶朎乮兛兝T嵶朎乯偺TCR偼丄兛嵔乮V兛丄J兛乯偲兝嵔乮J兝丄D兝丄V兝乯偱峔惉偝傟傞乮TCR2乯丅
丂兛嵔傕兝嵔傕丄峈尨偲寢崌偡傞柶塽僌儘僽儕儞偲摨條偵丄壜曄椞堟乮V椞堟乯偲僐儞僗僞儞僩椞堟乮C椞堟乯偐傜峔惉偝傟偰偄傞丅TCR傪僐乕僪偡傞堚揱巕偼丄柶塽僌儘僽儕儞偺堚揱巕偲摨條偵丄嵞峔惉乮gene rearrangement乯偝傟丄懡條側峈尨偵摿堎揑偵斀墳偡傞丅丂
丂嫻態偱偼丄帺屓斀墳惈T嵶朎傪丄彍嫀偡傞丗嫻態偵傗偭偰棃偨丄枹暘壔側T嵶朎偼丄慖戰傪庴偗丄嫻態偺峈尨採帵嵶朎乮HLA亄帺屓峈尨乯偲丄揔愗側恊榓惈傪桳偡傞嵶朎乮僋儘乕儞乯偼丄暘壔丒惉弉偡傞丅嫻態偺峈尨採帵嵶朎乮HLA亄帺屓峈尨乯偲嫮偔斀墳偡傞嵶朎乮恊榓惈偑崅偄僋儘乕儞丗帺屓斀墳惈偑嫮偄僋儘乕儞乯傗丄斀墳偑庛偄嵶朎乮恊榓惈偑掅偄僋儘乕儞乯偼丄巰柵偝偣傜傟傞丅枹暘壔側T嵶朎偺撪丄枹暘壔側T嵶朎偺98%埲忋偼丄巰柵乮傾億僩乕僔僗乯偝偣傜傟傞丅
丂嫻態偱丄兛嵔偲兝嵔傪峔惉偟偨T嵶朎偼丄枛徑寣丄儕儞僷愡丄銪憻側偳偵暘晍偡傞丅
丂兞嵔偲兟嵔偱峔惉偝傟傞TCR乮TCR1乯傪桳偡傞T嵶朎傕丄懚嵼偡傞丅偙偺T兞兟嵶朎乮兞兟T嵶朎乯偼丄枛徑寣傗儕儞僷愡偱偼丄儕儞僷媴偺1乣5亾偟偐懚嵼偟側偄偑丄旂晢傗彫挵偺忋旂傗擲枌偱偼丄T嵶朎偺50亾埲忋傪愯傔傞丅T兞兟嵶朎傕CD16傪敪尰偡傞傕偺傕懡偔丄MHC峉懇惈傕尒傜傟偢丄NK嵶朎條丄ADCC條偺嵶朎婡擻傪帩偪摼傞丅T兞兟嵶朎偼丄Th1嵶朎傊偺暘壔傪梷惂偡傞偲峫偊傜傟偰偄傞丅兞兟T嵶朎乮T兞兟嵶朎乯偼丄庡偵丄娞椶摯撪偱暘壔偡傞丅
丂嫻態奜暘壔T嵶朎乮T兞兟嵶朎乯偼丄NK嵶朎偲摨條偵丄Cd56丄CD57偑梲惈丅
丂嫻態偼丄偊傜屇媧忋旂壓偵偁傞儕儞僷媴廤崌偑敪払偟偨丅
丂偦傕偦傕丄儕儞僷媴偼丄儅僋儘僼傽乕僕乮扨媴乯偐傜恑壔偟偰棃偨偲峫偊傜傟偰偄傞丅
丂侾丏儅僋儘僼傽乕僕乮扨媴乯仺
丂俀丏NK嵶朎乮CD2+CD3-TCR-丄CD161+丄IL-2R兝+LFA-1+偺LGL乯仺
丂俁丏NKT嵶朎乮CD3+丄TCR偼堦庬椶+丄CD161+乯仺
丂係丏兞兟T嵶朎乮T兞兟嵶朎丗嫻態奜暘壔T嵶朎丗CD2+CD3+TCR+偱IL-2R兝+LFA-1+偺LGL乯仺
丂俆丏兛兝T嵶朎乮嫻態桼棃T嵶朎丗CD2+CD3+TCR+偱IL-2R兝-LFA-1-丗IL-2R兝傗LFA-1偼丄昁梫帪偵偺傒敪尰偡傞乯
丂偲暘壔偟偨偲峫偊傜傟傞丅
丂拲係丗昞柺偵IgM丄IgD丄CD40梲惈偺B嵶朎偼丄CD4丄CD154梲惈偺Th1嵶朎偲斀墳偡傞乮峈尨傪丄B嵶朎偐傜丄僿儖僷乕T嵶朎偺TCR丒CD3暋崌懱偵丄採帵偝傟傞乯丅
丂僿儖僷乕T嵶朎偐傜丄IL-4偑嶻惗偝傟傞偲丄B嵶朎偼丄IgG1傗IgE傪嶻惗偡傞傛偆偵側傞丅僿儖僷乕T嵶朎偐傜丄IFN-兞偑嶻惗偝傟傞偲丄IgG2a傗IgG3偑嶻惗偝傟傞傛偆偵側傞丅僿儖僷乕T嵶朎偐傜丄TGF-兝偑嶻惗偝傟傞偲丄IgG2a傗IgA偑嶻惗偝傟傞傛偆偵側傞丅
丂拲俆丗儕儞僷媴乮T嵶朎丄B嵶朎丄NK嵶朎側偳乯丄扨媴偼丄枛徑寣偺揾枟昗杮偱偼丄娵偄宍傪偟偰偄傞偑丄幚嵺偵偼丄儕儞僷媴傗扨媴偵偼丄摢晹偲壓晹乮旜晹乯偑懚嵼偟丄僾儗乕僩撪偱攟梴偡傞偲丄恾偺傛偆偵丄撍婲傪弌偟偨傝偟偰丄嬌嵼偟偨巔傪帵偡丅
丂拲俇丗墛徢惈僒僀僩僇僀儞偲偟偰偼丄TNF-兛丄IL-1兝丄IL-6丄IFN兞丄IL-8偑懚嵼偡傞丅墛徢惈僒僀僩僇僀儞偼丄妶惈壔儅僋儘僼傽乕僕傗妶惈壔寣娗撪旂嵶朎偐傜丄嶻惗偝傟傞丅
丂峈墛徢惈僒僀僩僇僀儞乮梷惂惈僒僀僩僇僀儞乯偲偟偰偼丄IL-4丄IL-10丄TGF-兝偑懚嵼偟丄妶惈壔儅僋儘僼傽乕僕側偳偐傜丄嶻惗偝傟傞丅側偍丄妶惈壔儅僋儘僼傽乕僕偼丄峈墛徢嶌梡偺偁傞僾儘僗僞僌儔儞僕儞偺PGE2傪傕丄嶻惗偡傞丅
丂T嵶朎僒僀僩僇僀儞偲偟偰偼丄Th1嵶朎乮TH1乯偐傜丄姶愼徢偺嵺偵丄IL-2丄IFN兞偑丄嶻惗偝傟丄Th2嵶朎乮TH2乯偐傜丄傾儗儖僊乕偺嵺偵丄IL-4丄IL-5偑丄嶻惗偝傟傞丅丂丂
丂拲俈丗Toll-like receptor 乮TLRs乯偼丄儅僋儘僼傽乕僕乮M冇乯傗庽忬嵶朎偵懚嵼偟丄昦尨懱乮嵶嬠傗僂僀儖僗側偳乯偵摿堎揑側抈敀丄DNA丄RNA傪擣幆偡傞丅偦偺寢壥丄嵶朎撪僔僌僫儖揱払宱楬偑妶惈壔偝傟丄墛徢惈僒僀僩僇僀儞傗丄僀儞僞乕僼僃儘儞偑丄嶻惗偝傟傞丅Toll偼丄僴僄乮攬乯偺堚揱巕偱丄恀嬠偺怤擖傪嶡抦偟偰丄妶惈壔僔僌僫儖傪揱払偟丄姶愼杊屼斀墳傪桿摫偡傞丅
丂Toll-like receptor 乮TLRs乯偺懠偵丄嵶朎昞柺偵偼丄曗懱C3儗僙僾僞乕丄Mannose receptor丄Scavenger receptor側偳偑丄懚嵼偟丄怤擖偟偨昦尨懱乮嵶嬠傗僂僀儖僗側偳乯偺怤擖傪丄擣幆偡傞丅
丂Mannose receptor偼丄嵶朎昞柺偺儗僋僠儞偱偁傝丄昦尨懱昞柺乮昞憌乯偺摐乮摐嵔偺儅儞僲乕僗乯偲寢崌偡傞丗Mannose receptor偼丄嵶嬠丄恀嬠丄僂僀儖僗姶愼嵶朎丄婑惗拵偺昞柺偺峈尨乮摐嵔偺儅儞僲乕僗乯偲丄寢崌偡傞丅
丂Scavenger receptor偼丄儕億抈敀傗丄儕億懡摐乮LPS乯側偳嵶嬠偺昞柺偵懚嵼偡傞摐帀幙偲傕丄寢崌偡傞丅
丂儅僋儘僼傽乕僕乮M冇乯昞柺側偳偵懚嵼偡傞丄曗懱C3儗僙僾僞乕丄Scavenger receptor丄Fc儗僙僾僞乕丄儗僋僠儞側偳偵傛傝丄怤擖偟偨昦尨懱乮嵶嬠傗僂僀儖僗乯偼丄婷怘偝傟丄僋儕傾儔儞僗偝傟傞丅
丂昦尨懱偺嬠懱枌惉暘乮LPS側偳乯偼丄嵶朎昞柺偵懚嵼偡傞TLR4/MD-2丄TLR2/TLR1丄TLR2/TLR6偵傛偭偰丄擣幆偝傟傞丅
丂僂僀儖僗桼棃偺DNA丄RNA偼丄嵶朎撪梓棻偵懚嵼偡傞TLR3丄TLR7丄TLR8丄TLR9偵傛偭偰丄擣幆偝傟傞丅
丂TRL乮Toll條庴梕懱乯偵偼丄TLR1乣TLR10偺懚嵼偑抦傜傟偰偄傞偑丄TLR8偲TLR10偺婡擻偼丄敾柧偟偰偄側偄丅TLR1丄TLR2丄TLR4丄TLR5丄TLR6偼丄嵶朎昞柺偵懚嵼偟丄TLR3丄TLR俈丄TLR9偼丄嵶朎幙撪偺僄儞僪僜乕儉偵懚嵼偡傞丅
丂TLR1偼丄TLR2偲丄僿僥儘擇検懱傪宍惉偟丄儁僾僠僪僌儕僇儞傗丄嵶嬠桼棃偺儕億儁僾僠僪乮僩儕傾僔儖儕億儁僾僠僪乯傪丄擣幆偡傞丅
丂TLR2偼丄TLR1傗丄TLR6偲丄僿僥儘擇検懱傪宍惉偡傞丅
丂TLR3偼丄dsRNA乮擇杮嵔RNA丗僂僀儖僗桼棃乯傪丄擣幆偡傞丅
丂TLR4偼丄MD2偲夛崌偟偰暋崌懱傪宍惉偟偰丄LPS乮僄儞僪僩僉僔儞乯丄儕億僞僀僐巁丄儕僺僢僪A傪丄擣幆偡傞丅LPS偼丄寣惔拞偱偼丄LBP乮LPS binding protein丗娞憻偱惗惉偝傟傞乯偲丄LPS/LPB暋崌懱傪宍惉偡傞丅TLR4丄MD-2丄CD14偼丄LPS儗僙僾僞乕暋崌懱傪宍惉偟丄LPS/LPB暋崌懱傪擣幆乮庴梕乯偡傞丅TLR4偼丄儅僋儘僼傽乕僕埲奜偵丄挵擲枌忋旂嵶朎丄堓擲枌忋旂嵶朎偵傕敪尰偟偰偄傞丅僺儘儕嬠乮H. pylori乯偐傜暘斿偝傟傞儕億懡摐椶乮LPS乯偼丄TLR4偵寢崌偟丄僒僀僩僇僀儞偺IL-8嶻惗傪桿摫偟丄堓擲枌偺墛徢乮堓墛乯偑婲偙傞丅
丂TLR5偼丄僼儔僕僃儕儞乮嵶嬠偺曏栄偵娷傑傟傞乯傪丄擣幆偡傞丅TLR5偼丄挵娗偺忋旂嵶朎偺栄嵶寣娗懁偵懚嵼偡傞丅惗懱偼丄挵娗偺忋旂嵶朎昞柺偵懚嵼偡傞挵撪嵶嬠乮忢嵼嵶嬠乯偵懳偟偰偼丄柶塽姲梕傪桿摫偡傞偑丄挵娗擲枌傪墇偊偰怤擖偟偨挵撪嵶嬠偵懳偟偰偼丄TLR5側偳偱丄擣幆偟乮姶抦偟乯丄柶塽墳摎傪偡傞丅
丂TLR6偼丄TLR2偲丄僿僥儘擇検懱傪宍惉偟丄儅僀僐僾儔僘儅桼棃偺儕億儁僾僠僪乮嵶嬠桼棃偺儕億儁僾僠僪偲偼丄傾僔儖婎偑堎側傞乯傪丄擣幆偡傞丅
丂TLR7偼丄ssRNA乮堦杮嵔RNA乯傗丄僀儈僟僝僉僲儕儞乮峈僂僀儖僗栻乯傪擣幆偡傞丅
丂TLR9偼丄旕儊僠儖壔CpGDNA乮嵶嬠偺DNA丗歁擕椶偺DNA偼丄C-G偺C偺晹暘偑丄儊僠儖壔偝傟偰偄傞乯傪丄擣幆偡傞丅僋儘儅僠儞峔憿偼丄DNA儊僠儖壔傗丄僸僗僩儞廋忺偵傛傝丄揮幨妶惈偑挷愡偝傟傞乮僄僺僕僃僱儕僢僋惂屼乯丅CpG乮僔僩僔儞-僌傾僯儞乯偼丄歁擕椶偱偼丄儊僠儖壔偑嵟傕崅昿搙偵擣傔傜傟傞僕僰僋儗僆僠僪丅CpA乮僔僩僔儞-傾僨僯儞乯傗丄CpT乮僔僩僔儞-僠儈儞乯巆婎傕丄儊僠儖壔偑擣傔傜傟傞丅
丂嶲峫暥專
丂丒媨嶁怣擵丄懠丗傢偐傝傗偡偄柶塽幘姵丂擔杮堛巘夛嶨帍丂摿暿崋乮侾乯丂惗奤嫵堢僔儕乕僘亅俇俈丄2005擭丏
丂丒扟岥崕丄懠丗昗弨柶塽妛乮戞俀斉丄堛妛彂堾丄2004擭乯丏
丂丒嶳杮堦旻丄懠丗僇儔乕恾夝丂恱懷偺惓忢峔憿偲婡擻丂俬倁丂寣塼丒柶塽丒撪暘斿丂乮擔杮堛帠怴曬幮丄2002擭乯丏
丂丒怷杮婔晇丗T儕儞僷媴偺柶塽偵偍偗傞栶妱丂擔杮堛巘夛嶨帍乮摿廤丂椪彴堛偺偨傔偺柶塽妛乯丂戞123姫丒戞11崋丄1733-1739, 2000擭丏
丂丒媨嶁怣擵丗僫僠儏儔儖僉儔乕嵶朎偺摥偒丂擔杮堛巘夛嶨帍乮摿廤丂椪彴堛偺偨傔偺柶塽妛乯丂戞123姫丒戞11崋丄1740-1742, 2000擭丏
丂丒嶰怷宱悽丗帺屓柶塽敪徢偺儊僇僯僘儉丂擔杮堛巘夛嶨帍乮摿廤丂椪彴堛偺偨傔偺柶塽妛乯丂戞123姫丒戞11崋丄1747-1752, 2000擭丏
丂丒娸杮拤嶰丄懠丗娾攇島嵗丂柶塽壢妛俁丂柶塽扴摉嵶朎乮娾攇彂揦丄1986擭乯丏
丂丒嬥揷堦岶丄奯惗墍巕丄懠丗怴柶塽妛憄彂侾侽丂僉儔乕嵶朎乮堛妛彂堾丄1983擭乯丏
丂丒嶳懞梇堦丄懠丗柶塽妛係丂嵶朎惈柶塽丂傾儗儖僊乕乮拞嶳彂揦丄1982擭乯丏
丂丒彫椦搊丗椪彴堛偺偨傔偺柶塽壢妛丂擔杮堛巘夛嶨帍丂惗奤嫵堢僔儕乕僘亅俆丂戞93姫丒戞8崋丄徍榓60擭丏
丂丒抾撪嬑丗憤愢丂CD45暘巕偲柶塽墳摎丂擔杮椪彴柶塽妛夛嶨帍丂Vol.15 No.1丄1-12丄1992擭丏丂丒埨曐揙丗NK-LAK嵶朎偺嵟嬤偺抦尒丂擔杮椪彴柶塽妛夛嶨帍丂Vol.15 No.6丄517-521丄1992擭丏
丂丒嶰塝憤堦榊丄懠丗挵儕儞僷憰抲乮GALT乯亅宍懺偲挷愡婡峔亅丂擔杮椪彴柶塽妛夛嶨帍丂Vol.15 No.6丄553-558丄1992擭丏
丂丒嶳忛梇堦榊丗憤墤丂徚壔娗偼柶塽憻婍偺侾偮偱偁傞丂擔杮彫帣壢妛夛嶨帍丂98姫4崋丄841-844丄1994擭丏
丂丒嶁堜寶梇丄壨尨崕梇丗僇儔乕恾夝丂恖懱偺惓忢峔憿偲婡擻丂俬俬俬徚壔娗丄擔杮堛帠怴曬幮丄2003擭丏
丂丒怴扟峅幚丗堓挵偼岅傞亅堓憡挵憡偐傜傒偨寬峃丒挿庻朄丄峅暥摪乮暯惉10擭弶斉丄暯惉12擭11嶞乯丏
丂丒栘壓朏堦丄抾岥婭峎丗僾儘僩儞億儞僾扵媮僔儕乕僘丄H.pylori姶愼偺桳柍偲堓巁暘斿摦懺丄僄乕僓僀姅幃夛幮丄CODE PT乮1乯369乮2005擭8寧嶌惉乯丏
丂丒嵵摗攷媣丗戞109夞擔杮彫帣壢妛夛妛弍廤夛丂暘栰暿僔儞億僕僂儉丗擕梒帣婥娗巟歜懅帯椕偺憗婜夘擖丂傾儗儖僊乕惈墛徢惈幘姵偵娭傢傞堚揱揑攚宨丄僄僺僕僃僱僥傿僢僋惂屼偲娐嫬場巕丄擔杮彫帣壢妛夛嶨帍丄111姫1崋丄10-15暸乮2007擭乯丏