NK細胞
NK細胞は、生体内で、ウイルス感染細胞や、一部の腫瘍細胞を認識し、傷害する。
NK細胞の標的細胞障害作用は、抗原特異性や、MHC拘束性はなく、以前に出会ったことがない標的細胞も傷害する(自然免疫)。
NK細胞は、特有の糖鎖を表面に有する細胞、MHCクラスI分子を失った細胞を、標的にして、傷害する。
NK細胞は、NK細胞受容体(CD161)により、標的細胞の特定の糖鎖を認識し、また、MHCクラスI分子受容体により、標的細胞のMHCクラスI分子(の有無)を認識し、MHCクラスI分子を失った細胞を、障害する。
MHCクラスI分子受容体は、標的細胞が、MHCクラスI分子を発現している際には、抑制型のシグナルを伝達し、NK細胞は、標的細胞を、障害しない。しかし、MHCクラスI分子受容体は、標的細胞が、MHCクラスI分子を発現していない際には、活性化型のシグナルを伝達し、NK細胞は、標的細胞を、障害する。
NK細胞は、細胞膜表面に、糖脂質のアシアロGM1を有している。また、NK細胞は、細胞膜表面のCD161により、標的細胞の細胞膜に存在する特定の糖鎖構造(アシアロGM2など)と、結合する。
NK細胞活性は、脂肪摂取を制限すると、増強する:NK細胞活性は、高脂肪食(特に、n-3系の多価不飽和脂肪酸)を摂取すると、低下する。
NK細胞は、表面マーカーは、CD2+CD3-TCR-、CD161+、IL-2Rβ+LFA-1+で、形態は、大型顆粒リンパ球(LGL)。
1.NK細胞の標的細胞障害
NK細胞(natural killer cells:ナチュラルキラー細胞)は、生体内で、ウイルス感染細胞や、一部の腫瘍細胞を認識し(結合し)、傷害する。
NK細胞は、TCR(T細胞レセプター)が発現しておらず、T細胞ではない。
腫瘍細胞を傷害する活性は、キラーT細胞(CTL:cytotoxic T cells)の方が強い。
NK細胞の標的細胞障害作用は、以下の点で、キラーT細胞と異なっている。
・抗原特異性はない:感作を必要とせず、以前に出会ったことがない標的細胞も傷害する(自然免疫)。
・MHC(主要組織適合抗原)により拘束されない(MHC非拘束):NK細胞は、K562(MHCを保有していない)などの、他人由来の白血病細胞も傷害する。K562は、MHCを保有していないが、CD54(ICAM-1)、CD58(LFA-3)が細胞表面上に存在し、それぞれ、NK細胞表面のLFA-1(CD11a)、LFA-2(CD2)と、接着する。
なお、ウイルス感染細胞を傷害するキラーT細胞は、自己と同じMHCクラスI分子を発現したウイルス感染細胞しか、傷害しない:キラーT細胞は、抗原が同じでも、MHCクラスI分子が自己と異なる細胞は障害しない。これを「MHC拘束」と呼ぶ。
反対に、NK細胞は、MHCクラスI分子を発現した細胞に対しては、その傷害作用が、抑制される。NK細胞は、標的細胞が、MHCクラスI分子の発現が弱い程、強い、細胞障害活性を示す。CMV(サイトメガロウイルス)は、ゲノム中に、ヒトMHCクラスI分子と相同性の高い遺伝子を有していて、感染した細胞に発現させ、NK細胞からの攻撃(障害)を免れる。
NK細胞は、ウイルス感染の初期から働く。
ウイルス感染時に、ウイルス感染細胞から産生される、IFN-α(注1)、IFN-βは、NK細胞活性を増強させる。
NK細胞活性が低下すると、かぜをひきやすくなる。
ひとつのNK細胞は、次々と別の種類の標的細胞を傷害出来る。
IL-12で活性化されたNK細胞は、IFN-γを産生する(注2)。IFN-γは、NK細胞活性を増強させる(注1)。
NK細胞は、MS(多発性硬化症)の寛解期には、IL-5を産生して、病態の悪化を阻止している可能性があるという。
NK細胞活性は、脂肪摂取を制限すると、増強する(脂肪摂取を全エネルギーの22%以下にする)。また、NK細胞活性は、高脂肪食、特に、n-3系の多価不飽和脂肪酸を多く含む魚油を摂取すると、低下する。
入浴などで、体温が1℃上昇すると、NK細胞活性が、増加する。体温が低下すると、免疫力(NK細胞活性)が、37%低下する。
2.NK細胞の形態
NK細胞は、細胞質に富んだ、大型のリンパ球で、細胞質にアズール好性の粗大顆粒を持つ。 NK細胞の核は、腎臓のような型をしている。
NK細胞は、形態的には、大型顆粒リンパ球(large granular lymphocyte:LGL):LGLは、細胞内顆粒により、細胞障害機能を示すリンパ球である。一般的に、LGLとは、NK細胞を意味するが、活性化されたCTL(キラーT細胞)や、Tγδ細胞(γδT細胞:γδ型TCR発現するT細胞)も、LGLの形態を示す。LGLは、正常人の末梢血中リンパ球の1〜8%を占める。NK細胞は、表面マーカーは、CD2+CD3-TCR-だが、LGLの中には、胸腺外分化T細胞が存在し、その表面マーカーは、CD2+CD3+TCR+(intermediate:ヒトではbright)。なお、胸腺で分化した胸腺由来T細胞も、その表面マーカーは、CD2+CD3+TCR+(bright)。
NK細胞は、末梢血や、脾臓(マウス)のリンパ球の、3〜10%を占める。
NK細胞は、末梢血中(循環血中)のみならず、肝類洞、腸管上皮、肺上皮、胎盤などの組織中にも、存在する。
3.NK細胞の表面抗原
ヒトNK細胞の表面抗原の特徴は、下記の如く。
NK細胞は、CD2+CD3-TCR-で、IL-2Rβ+LFA-1+(常時発現)のLGL。
・NK細胞は、T細胞とも、B細胞とも、表面抗原(表面マーカー)が、異なる:NK細胞は、T細胞特異的マーカー(TCR、CD3、CD4、CD8)、B細胞特異的マーカー(Ig、CD19、CD20)を発現しない。
・CD2陽性:CD2(LFA-2)は、羊の赤血球(E:SRBC)と結合する際の、E-ロゼット受容体。T細胞は、ノイラミニダーゼでシアル酸を処理した、羊の赤血球と、ロゼットを形成する。NK細胞は、T細胞と同様に、羊の赤血球とロゼットを形成する(E-rosette forming cells、注3)。なお、抗原提示細胞は、CD58(LFA-3)と言う、CD2と結合するリガンドを有している。
・CD3陰性(CD3の構成成分のζ鎖ダイマーは持つ)←T細胞は、細胞表面のCD3陽性
・CD4陰性
・CD16陽性:CD16は、免疫グロブリンのFc部分に対するリセプター(FcγRIII:Fcγ受容体タイプIII)
・CD56陽性:CD56は、NCAM-1
・IL-2レセプター陽性:NK細胞は、低親和性のIL-2レセプターを持つ(注1)。高濃度のIL-2で活性化され増殖したのが、LAK細胞(lymphokine-activated killer cells)
LAK細胞の前駆細胞となるのは、IL-2Rβを常時発現している(休止状態でも発現している)NK細胞と、胸腺外分化T細胞(γδT細胞)。なお、胸腺由来T細胞も、通常は、IL-2Rα、IL-2Rβを発現していない(陰性)だが、抗原やIL-2により活性化されると、IL-2Rα、IL-2Rβを発現する。従って、胸腺由来T細胞も、LAK細胞の前駆細胞となる。NKT細胞(CD244陽性)も、LAK細胞の前駆細胞となると思われる。
・TCR(T細胞レセプター)陰性←NK細胞は、T細胞と異なる
・免疫グロブリン陰性←B細胞は、表面免疫グロブリン(sIg)陽性
・アシアロGM1:抗アシアロGM1抗体をマウスに投与すると、リンパ球系組織のすべてからアシアロGM1陽性細胞が消失し、NK細胞活性も除去される。これは、NK細胞が、細胞膜表面に、糖脂質のアシアロGM1を有している(NK-I、NK-Tなど)ことと、関連している。アシアロGM1は、ガングリオシド(シアル酸を有する酸性糖脂質)のGM1(注4)からシアル酸を除いた中性糖脂質であり、細菌や細菌毒素と結合するという。アシアロGM1は、マウスのマクロファージにも発現されている。標的細胞が、アシアロGM2、グロボシドなど、特定の糖脂質を、細胞膜へ発現しなくなると、NK細胞により、障害(攻撃)されにくくなる(NK抵抗性になり、NK細胞活性が低下する)。これは、NK細胞が、CD161(レクチン様受容体)により、標的細胞の細胞膜に存在する特定の糖鎖構造(GalNAcβ1-4Galβ1-4Glc糖鎖:アシアロGM2など)を、認識することと関連している。
・未熟なT細胞も、NK細胞と同様に、アシアロGM1を細胞表面に発現する。マウス胎仔での研究結果によると、胎齢12日頃(胸腺原基に、初めて幹細胞が出現する)、胸腺内のリンパ球系細胞は、ほとんど全て(95%)、アシアロGM1を細胞表面に発現している。しかし、胎齢と共に、Thy-1、Lyt-1、Lyt-2を発現するリンパ球が増加し、生下時(胎齢20日)には、アシアロGM1を細胞表面に発現するリンパ球は、胸腺内に、ほとんど存在しない(3%以下)。
・NK細胞は、ヌードマウスや、胸腺摘出マウスにも、存在する。
・NK細胞は、免疫グロブリン・ファミリー(イムノグロブリンスーパーファミリー)に属するCD2、CD16(FcγRIII)、CD56(NCAM-1)、CD57、インテグリン・ファミリーに属するLFA-1、VLA-4、アニマルレクチン・スーパーファミリーに属するLy-49、NKG2、NKR-P1(CD161)が、陽性
・NK細胞は、CD244陽性。CD244は、NK細胞、NKT細胞、LAK細胞が陽性
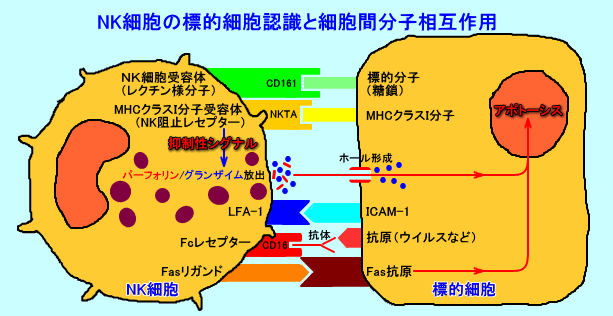
4.NK細胞の標的細胞認識と細胞間分子相互作用
NK細胞は、NK細胞受容体(CD161:NKR-P1)により、標的細胞の特定の糖鎖を認識し、MHCクラスI分子受容体により、標的細胞のMHCクラスI分子(の有無)を認識し、MHCクラスI分子を失った細胞を、障害する。
NK細胞のMHCクラスI分子受容体には、KIR受容体(免疫グロブリン・ファミリー)と、NKG2受容体(C型レクチン・ファミリー)とが、存在する。両者共に、NK細胞活性を亢進させる活性化型と、NK細胞活性を抑制する抑制型の、2種類が存在する。
NK細胞は、標的細胞の糖鎖を認識したNK細胞受容体から、「オン」のシグナルを受け、抑制型のMHCクラスI分子受容体から、「オフ」のシグナルを受けない時(活性型のMHCクラスI分子受容体から、「オン」のシグナルを受けた時)、標的細胞を、障害する(破壊する)。
NK細胞は、
・特有の糖鎖を表面に有する細胞
・MHCクラスI分子を失った細胞
を、標的にして、傷害する。
NK細胞表面には、レクチン様分子のNK細胞受容体(NKレセプター)が存在し、標的細胞の糖鎖を認識する(結合する)ものと考えられて来た(レクチンとは、植物などに存在する蛋白質で、細胞表面の特定の糖鎖構造と結合する性質がある)。
NK細胞は、NK細胞受容体(NKレセプター)を用いて、特有の糖鎖(GalNAcβ1-4Galβ1-4Glc糖鎖、1,3galactosyl epitope?)を表面に有する標的細胞に、結合するものと考えられる。
キラーT細胞は、TCR・CD3複合体により、抗原提示細胞(抗原呈示細胞)のMHCクラスI分子+抗原ペプチドを、認識するのに対して、NK細胞は、NK細胞受容体(NKレセプター)により、標的細胞の糖鎖を、認識する。
a.CD161
CD161(NKR-P1:NK cell receptor-P1、NK1.1も一員)は、NK細胞(CD3陰性)と、一部のT細胞(CD3陽性のNKT細胞、CD4陽性やCD8陽性のT細胞)、活性化された単球に発現している。
CD161は、レクチン様受容体(2型の膜貫通型C-型レクチンでCa2+依存性)だが、何がリガンドなのかは、知られていない。NK細胞は、CD161(レクチン様受容体)により、標的細胞の細胞膜に存在する特定の糖鎖構造を認識すると推定されている。特定の糖鎖構造としては、GalNAcβ1-4Galβ1-4Glc糖鎖(アシアロGM2など)を有するプロテオグリカンや、糖脂質(ガングリオシド)、糖蛋白などが、標的構造と推定されている。
CD161(NKR-P1)は、Fcレセプターのγサブユニット(FcRγ鎖)と会合して活性化シグナルを伝達する。
α-(1-3)-Dグルカンは、CD161(NKR-P1)を刺激し、NK細胞やNKT細胞を活性化させる。
なお、β-(1-3)-Dグルカンは、IL-12の産生を誘導し、マクロファージを活性化する。β-(1-6)-Dグルカンは、NK細胞、T細胞、及び、マクロファージを活性化すると考えられている(注5)。
2).MHCクラスI分子受容体
NK細胞は、MHCクラスI分子と結合するMHCクラスI分子受容体(MHCクラスI分子を認識する受容体)を、有している。
MHCクラスI分子受容体は、標的細胞が、MHCクラスI分子を発現している際には、抑制型のシグナル(「オフ」のシグナル)を伝達し、NK細胞は、標的細胞を、障害しない。しかし、MHCクラスI分子受容体は、標的細胞が、MHCクラスI分子を発現していない際には、活性化型のシグナル(「オン」のシグナル)を伝達し、NK細胞は、標的細胞を、障害する。
NK細胞表面のMHCクラスI分子受容体には、KIR(killer inhibitory receptor:免疫グロブリン・ファミリー)と、NKG2(C型レクチン・ファミリー)とが、存在する。NKG2受容体としては、ヒトではCD94/NKG2が、マウスではLy-49が知られている。
これらのMHCクラスI分子受容体は、典型的なMHCクラスI分子(HLA-A、B、C)以外に、MHCクラスI相同分子群(HLA-E、MICA、MICBなど)をも、認識する(結合する)。
NK細胞は、標的細胞が、MHCクラスI分子の発現が弱い程、強い、細胞障害活性を示す。
NK細胞のMHCクラスI分子受容体には、KIR受容体(免疫グロブリン・ファミリー)と、NKG2受容体(C型レクチン・ファミリー)とが、存在する。
両者のMHCクラスI分子受容体は、共に、NK細胞活性を亢進させる活性化型と、NK細胞活性を抑制する抑制型の、2種類が存在する。活性型は、ITAM(immunoreceptor tyrosine-based activating motif)構造を有するシグナル伝達分子が会合している。抑制型は、ITIM(immunoreceptor tyrosine-based inhibitory motif)構造をしている。
正常細胞は、MHCクラスI分子を発現しているので、抑制型のMHCクラスI分子受容体から、「オフ」のシグナルが優位に発せられ、NK細胞は、障害しない。癌化した細胞や、ウイルス感染細胞は、MHCクラスI分子の発現が、減弱すると、活性化のMHCクラスI分子受容体から、「オン」のシグナルが優位に発せられ、NK細胞は、障害する。
このように、MHCクラスI分子受容体の抑制型は、NK細胞の障害活性を阻止する、NK阻止レセプター(KIR:killer inhibitory receptor)として、機能する。また、MHCクラスI分子受容体の活性化型は、NK細胞の障害活性を活性化する、NK活性化レセプター(KAR:killer activation receptor)として、機能する。
KIRは、T細胞も発現している。
NK細胞は、標的細胞の糖鎖を認識したNK細胞受容体から、「オン」のシグナルを受け、抑制型のMHCクラスI分子受容体から、「オフ」のシグナルを受けない時(活性型のMHCクラスI分子受容体から、「オン」のシグナルを受けた時)、標的細胞を、障害する(破壊する)。
NK細胞表面に存在する、NKB1などの分子は、MHCクラスI分子受容体であり、標的細胞表面の特定のMHCクラスI分子と結合する(NKB1は、T細胞表面にも存在するMHCレセプター)。
このようにして、NK細胞が、抑制性レセプター(抑制受容体)のNK阻止レセプターにより、標的細胞表面に、自己のMHCクラスI分子(α鎖)を認識すると、抑制性シグナルが、NK細胞内に伝達され、チロシンリン酸化が阻害され、NK細胞の活性化が阻止される(注6)。
そのため、NK細胞は、MHCクラスI分子を発現している自己の細胞を、傷害しにくい(自己寛容)。
そして、MHCクラスI分子を失ったウイルス感染細胞や腫瘍細胞などは、NK細胞により傷害されやすい。
このことは、MHCクラスI分子を介して分子を認識し、活性化する細胞傷害性T細胞(cytotoxic T lymphocytes:CTL、CD8陽性のキラーT細胞)と異なる。
a.NKTA
KIRは、免疫グロブリン・ファミリーに属し、ヒトでは、NKTAの存在が知られている。
b.CD158
NK細胞表面のCD158は、KIRとして、標的細胞のMHCクラスI分子(HLA-C)を認識し、NK細胞に傷害作用を抑制するように、抑制性シグナルを伝える。
c.NKG2D
NKG2Dは、C型レクチン様受容体ファミリー(Ca2+依存性)に属するレセプターであり、すべてのCD8陽性T細胞、NK細胞、Tγδ細胞(γδT細胞)に発現が認められる。
ヒトでは、MICA、MICBと言ったnon-classicalなMHCクラスI様蛋白質を、正常細胞は発現していないが、腫瘍細胞は発現している。
NK細胞や、Tγδ細胞(γδT細胞)では、NKG2D(レセプター)で、MICAやMICB(リガンド)を認識して、細胞障害活性を示す。NKG2D(レセプター)は、C型レクチン様受容体ファミリ、DAP10(アダプター)と複合体を形成して、シグナルを伝達する:DAP10を介して、PI-3キナーゼのP85サブユニットが活性化され、NK細胞の標的細胞障害が行われる。
NKG2D-DAP10複合体は、マウスにも存在し、RAE-1分子をリガンドとして認識する。
3).LFA-1
NK細胞表面には、接着分子のLFA-1(human leukocyte function-associated antigen-1)が存在する。
NK細胞が標的細胞と結合(接着)するには、LFA-1を介して、標的細胞表面のICAM-1との結合も必要。
LFA-1は、α鎖(CD11a)とβ鎖(CD18)で構成される。β鎖(CD18)は、補体レセプターのCR3のβ鎖と同じ。
LFA-1は、MAc-1、p150,95同様に、β2インテグリン群。
4).Fcレセプター
NK細胞表面には、免疫グロブリン(IgG)のFc部分と結合する、Fcレセプター(FcγRIII)が存在する。このFcレセプターの糖蛋白質は、CD16。
NK細胞は、Fcレセプターを介して、標的細胞表面に結合している、免疫グロブリン(抗体)のFc部分と結合し、標的細胞を傷害出来る。
このような、抗体を介する標的細胞の破壊は、ADCC(antibody-dependant cell-mediated cytotoxicity:抗体依存性細胞障害)と呼ばれ、単球・マクロファージも行う。
5).Fasリガンド
NK細胞は、細胞表面にFasリガンドを有しており、標的細胞表面のFas抗原(Fas受容体)と結合することで、標的細胞に(核クロマチンの濃縮、DNA断片化による細胞死)を引き起こす。
SLEなどの自己免疫疾患では、可溶性Fas抗原が増加しており、Fas抗原を発現した自己抗原反応性リンパ球が、アポトーシスにより除去されないことが、発症の原因とも言われる。
6).CD57(HNK-1糖鎖抗原)
マウスで、単クローン抗体HNK-1によって認識される抗原分子は、HNK-1糖鎖抗原と呼ばれる。
HNK-1糖鎖抗原は、ヒトのNK細胞などの細胞表面にも発現しており、別名CD57、Leu7と呼ばれている(単クローン抗体を開発したメーカーが異なるために、違う命名がされているが、CD57とLeu7は、同じ抗原を認識している)。
HNK-1糖鎖抗原は、N-アセチルラクトサミン構造の末端に、3位の硫酸化されたグルクロン酸が結合しているという。なお、N-アセチルラクトサミン(N-acetyllactosamine:LacNAc:Galβ1-4GlcNAc)は、ガラクトース(Gal)とN-アセチルグルコサミン(GlcNAc)とが、β-1,4結合したニ糖類。
HNK-1糖鎖抗原は、免疫系や神経系の細胞に分布している:HNK-1糖鎖抗原は、糖蛋白質や糖脂質に発現されている。糖蛋白質に関しては、細胞接着分子に発現している。
HNK-1糖鎖抗原は、神経系においては、細胞の接着や移動、神経突起の伸長に関与していると考えられている。
HNK-1糖鎖抗原は、多くの神経系細胞接着分子(免疫グロブリンスーパーファミリーやインテグリン・ファミリーに属する)に見出されているという。
5.NK細胞による標的細胞障害のメカニズム
NK細胞は、標的細胞に結合し、活性化されると、細胞内顆粒中のパーフォリン(perforin)を放出し、標的細胞の細胞膜の結合面に、穴(ホール)を開ける。パーフォリンは、補体第9成分と相同性を有する蛋白質で、NK細胞の顆粒内(酸性で、Ca2+濃度が低い)では、単量体として、貯蔵されている。
さらに、NK細胞顆粒から放出される、グランザイム(granzyme:セリンエステラーゼ)が、パーフォリンが開けた穴から、標的細胞に侵入し、標的細胞にアポトーシスを誘導し、標的細胞の核を断片化させて、標的細胞を傷害する。
NK細胞の顆粒内部は、酸性でCa2+濃度が低い為、パーフォリンは、単量体で存在する。NK細胞が活性化されると、NK細胞の顆粒膜が、NK細胞の細胞質膜と融合し、脱顆粒が起こり、パーフォリンやグランザイムが、NK細胞の外に放出される。
NK細胞から放出されたパーフォリンは、Ca2+(カルシウムイオン)存在下で、重合し、標的細胞の細胞膜に挿入されて、直径約160Åの穴(ホール)が、形成される。
NK細胞から放出されたグランザイムは、パーフォリンが形成した穴(ホール)を通過して、標的細胞に侵入して、標的細胞に、アポトーシスを誘導する。
Ca2+拮抗剤(トリフルオペラジン)は、NK細胞活性を抑制する。
なお、アスピリン、インドメタシンは、NK細胞活性を抑制しない。
NK細胞は、細胞表面にFcレセプター(CD16)を有しており、免疫グロブリン(抗体)と結合した標的細胞を、ADCCにより、傷害する。
NK細胞は、細胞表面にFasリガンドを有しており、Fas抗原(Fas受容体)を発現している標的細胞に結合して、アポトーシスを起こす。
その他、NK細胞は、NKCF(natural killer cytotoxic factor)も放出する。
6.NK細胞の産生するサイトカイン
NK細胞は、何種類ものサイトカインを産生する。
特に、NK細胞は、IFN-γ、TNF-αを産生する。これらの、サイトカインは、腫瘍細胞の増殖を抑制し、炎症を惹起する。また、IFN-γは、NK細胞活性を増強させる(ポジティブフィードバック)。
7.NK細胞を活性化させるサイトカイン
IFN-α、IFN-β(白血球、線維芽細胞が産生)、IFN-γ(T細胞、NK細胞が産生)、IL-2(T細胞、特に、Th1細胞が産生)、IL-15(広範な体細胞が産生)は、NK細胞の標的細胞障害活性(キラー活性)を、増強させる。
IL-2、IL-12(B細胞、単球、樹状細胞が産生する)、IL-15は、NK細胞の増殖を誘導する。IL-12は、IFN-γ産生を誘導する。
IL-15は、NK細胞の発生・分化に必須のサイトカイン。
NK細胞は、主要なインターフェロン産生細胞でもあり、IFN-α、IFN-β、IFN-γを産生する。
NK細胞は、ウイルス感染を感知する:NK細胞は、MHCクラスI分子受容体(KIR)により、MHCクラスI分子に結合したウイルスペプチドを認識して、活性化され、IFN-α、IFN-β、IFN-γや、TNF-αを、産生する。その結果、NK細胞は、単核食細胞(マクロファージ)、血管内皮細胞、好中球などを、活性化し、局所に炎症反応を起こす。
8.リンパ球の分化とNK細胞
リンパ球は、マクロファージ(単球)から進化したが、マクロファージ→NK細胞(CD2+CD3-TCR-、CD161+、CD7+CD56+、IL-2Rβ+LFA-1+のLGL)→NKT細胞(CD3+、TCRは一種類+、CD161+)→γδT細胞(胸腺外分化T細胞:CD2+CD3+TCRγ/δ+、IL-2Rβ+LFA-1+のLGL)→αβT細胞(胸腺由来T細胞:CD2+CD3+TCRα/β+、IL-2Rβ-LFA-1-:IL-2RβやLFA-1は、必要時にのみ発現する)の順に、分化したと考えられる。
NK細胞は、血管内皮細胞同様に、マクロファージから分化したので、細菌(黄色ブドウ球菌など)を貪食する性質がある。
9.NK細胞活性の測定
NK細胞活性は、標的細胞(K562細胞やRaji細胞)を51Crで標識(ラベル)し、検索する患者のリンパ球(単核球:Ficoll比重遠沈法で分離)と混合培養後、上清中に検出される51Crの放射能活性を測定する(51Cr遊離法)。
51Crをスピッツ内の遠沈させた標的細胞に滴下(100μCi)し、37℃で1時間放置(インキュベイト)し、標識(ラベル)し、その後、スピッツに培養液(RPMI1640液)を加え、3回、洗浄する。
51Crを標的細胞に滴下すると、51Crは、Cr6+イオンとして標的細胞の細胞膜を経て細胞質に入り、還元されてCr3+となり、細胞質分子と結合する。NK細胞によって、51Cr標識した標的細胞が破壊されると、細胞内から51Crが放出される(一度放出された51Crは、再度、生きている標的細胞に取り込まれない)。
マイクロプレートの小穴(well)で、患者の単核球(E:エフェクター細胞:4×105/well)と、51Cr標識標的細胞(T:Target cell:K562等:1×104/well)を混ぜ(E:T比=40:1、200μl/well)、37℃5時間放置(5%炭酸ガス培養器中)した後、マイクロプレート用の遠沈器で、1,000回転(rpm)で5分間遠沈し、上清を100μl採取し、ガンマ・カウンターで、51Cr放射能活性(cpm)を測定する(検体解離cpm)。
NK細胞活性(% cytotoxicity)=(検体解離cpm−自然解離cpm)÷(最大解離cpm−自然解離cpm)×100
最大解離cpm:標識標的細胞を凍結融解(3回)させたcpm
自然解離cpm:標識標的細胞(T)だけで、患者の単核球(E)を加えないwellのcpm
51Cr遊離法によるNK細胞活性の測定に用いられる標的細胞(培養株化細胞)には、K562(慢性骨髄性白血病由来)、MOLT-4(Tリンパ芽球細胞由来)、Raji(バーキットリンパ腫由来)が用いられた。
これらの標的細胞は、加熱不活化させた5%牛胎仔血清(fetal bovine serum:FCS)を加えたRPMI1640液で、炭酸ガス培養器で、培養を継続して、用意する。
10.その他
・ヨクイニンは、生薬で、イネ科ジュズダマ属ハトムギ(ハト麦)の種皮を除いた種仁。
ヨクイニンには、澱粉(52%)、蛋白質(18%)、脂肪油(7%)、グリカン類、benzoxazinones、脂溶成分のCoixenolideが含まれている。
ヨクイニンは、ヨクイニンエキスとしては、通常、成人、1.0〜2.0g/日を3回に分割して内服する。ヨクイニンエキス錠「コタロー」は、成人、9〜18錠/日を3回に分割して内服する。ヨクイニンエキス散「コタロー」は、成人、3.0〜6.0gを3回に分割して内服する(ヨクイニンエキス錠「コタロー」は、2g/1包を1日3包、内服)。
ヨクイニンエキス錠を服用した健常人は、末梢血白血球数や分画に有意な変動は見られないが、末梢血リンパ球サブセットで、NK細胞であるCD16+CD57-細胞や、MHC非拘束性細胞障害性T細胞であるCD3+CD56+細胞の比率が、有意に増加する。
ヨクイニンエキスは、尋常性疣贅や、青年性扁平疣贅に保険適応が承認されている。
ヨクイニンは、伝染性軟属腫にも有効。
注1: NK細胞活性は、IFN-αやIL-2やIFN-γと共に培養すると、増強する:
・ヒトNK細胞活性は、IFN-αと共に培養すると、活性の増強が培養1日以内に見られ、培養2日目以降は、活性が対照レベルに低下する。
・ヒトNK細胞活性は、IL-2と共に培養すると、活性の増強が培養2日目からピークになり、培養3日目も活性の増加が持続する。
・ヒトNK細胞活性は、IFN-γと共に培養すると、IFN-αやIL-2の活性増強より、遅れて増強し、培養3日目をピークに、活性が次第に対照レベルに低下する。
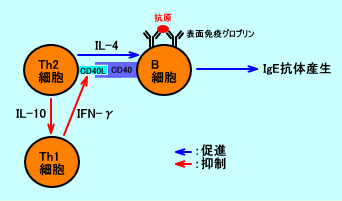
注2:IFN-γ(interferon-gamma)は、主に、CD4陽性ヘルパーT細胞(特に、Th1細胞)やCD8陽性キラーT細胞(CTL)が産生するが、IL-12で活性化されたNK細胞やNKT細胞も、IFN-γを産生する。なお、IL-12は、好中球、樹状細胞が産生する。
IFN-γは、抗ウイルス効果、マクロファージやNK細胞やCTLの細胞障害活性の増強作用がある。
IFN-γは、MHCクラスI分子、MHCクラスII分子、Fcγレセプター、TNFレセプターの発現を、増加させる。
Th1細胞が産生するIFN-γは、Th2細胞のCD40リガンド(CD40L、gp39)発現を抑制し、IgE抗体産生を抑制する。
注3:T細胞や、NK細胞は、羊の赤血球(E)を、細胞表面に結合させ、E-ロゼットを形成する。羊の赤血球(E)は、SRBC(sheep red blood cells)とも、略された。
E-ロゼットを形成させるには、予め、羊の赤血球を、ノイラミニダーゼ(neuraminidase)で処理して置く。ヘパリンを、血液1ml当たり50単位加えて、血液を採取し、試験管の中で、Ficoll-Hypaque液に重層させ、比重遠沈法(1,500〜20,00rpm、30分)にて、中間層の単核球層を採取する。単核球(MNC:mononuclear cells)層には、リンパ球、単球、血小板が含まれる。単核球層を、PBS(リン酸加生食、リン酸緩衝生理食塩水溶液)で、3回洗浄する。試験管内で、FCS(胎児ウシ血清)を10%加えたRPMI1640液(培養液)に、洗浄した単核球(MNC)を2×106/ml、ノイラミニダーゼ処理SRBC(E)を2×108/mlに浮遊させる(MNC:SRBC=1:50〜200)。良く攪拌し(ピペッティグする)、37℃15分間incubateする。その後、1,000rpmで5分間遠沈して、単核球とSRBCとを密着させた後、氷水中で、1時間、反応させる(ノイラミニダーゼで処理したSRBCを用いた場合、23℃でも良い)。その後、試験管の底面に遠沈した細胞(MNC+SRBC)を、静かに、ピペッティグにて、浮遊させる。浮遊液を、1滴、スライドガラスに落とし、カバーグラスをかけて、顕微鏡で観察し、ロゼットを形成した単核球の百分率を、求める。
モノクローナル抗体により、表面マーカー(表面抗原)の検査が行われるまでの時代は、白血病細胞などを分類する際、E-ロゼット形成細胞(E-rosette forming cells)をT細胞、細胞表面に蛍光抗体法で免疫グロブリンが存在する細胞をB細胞と、識別した。
CD2(LFA-2)は、羊の赤血球の受容体(E-ロゼットレセプター)。
NK細胞も、高率にCD2(LFA-2)を有していて、T細胞同様に、E-ロゼットを形成するが、低親和性と言われた。
日本人の多くは、EBウイルス(EBV)に対する抗体(EBV-VCA-IgG抗体など)が陽性で、体内に、EBウイルスに感染した細胞を有している。
E-ロゼット形成させた後、再度、Ficoll-Hypaque液に重層させ、比重遠沈すると、E-ロゼット形成細胞(T細胞)は、試験管底に沈み、中間層のnon-T細胞(≒B細胞)を採取する。
non-T細胞を、FCSを加えたRPMI1640液(培養液)にて、培養すると、EBウイルスにより形質転換された(transformed)B細胞株(Bリンパ芽球::lymphoblastoid cell lines:LCL)が、樹立されることが多い。このようなLCLは、癌細胞と同様に、試験管内では、無期限に増殖を繰り返すので、長期継代培養が可能なことが多いが、EBウイルスの種類によっては、他のウイルスのように細胞破壊的に作用し、長期継代培養出来ないこともある。
なお、CDは、cluster of differentiationの略。CD分類は、ヒト血液細胞上の分子(抗原)の、国際的な分類法になっている。
1970年代後半から、白血球表面の分子(抗原)に特異的に結合するモノクローナル抗体が、各社(各研究グループ)から、発売された。
モノクローナル抗体のLeuシリーズは、Becton Dickinson社が、開発した。OKTシリーズは、Ortho社が開発した。
しかし、各社により、異なる名称が付けられた為、これらのモノクローナル抗体を、国際的に分類して、統一した名称を付けるために、CD分類が、始まった。
注4:ガングリオシドは、シアル基や硫酸基を有する糖脂質。ガングリオシドは、神経系の修復(神経細胞の突起伸長)、細胞の分化・増殖、種々の物質(毒素、ウイルスなど)に対するレセプター機能、ホスホキナーゼ活性化などの整理活性を有する。
ガングリオシドのGM1は、神経系、特に、脳灰白質を中心に広く存在している。
ガングリオシドのGM1は、細胞内小器官のリソソーム内の酸性加水分解酵素(GM1βガラクトシダーゼ、GM2β-Nアセチルガラクトアミニナーゼ)により、糖鎖が順次分解され、GM1→GM2→GM3となる。
GM1:Galβ1-3GalNAcβ1-4[NeuAcα2-3]Galβ1-4Glcβ1-1Cer
GM2:GalNAcβ1-4[NeuAcα2-3]Galβ1-4Glcβ1-1Cer
GM3:NeuAcα2-3Galβ1-4Glcβ1-1Cer
なお、Gal:ガラクトース、GalNAc:N-アセチルガラクトサミン、NeuAc:N-アセチルノイラミン酸、Glc:グルコース、Cer:セラミド(Ceramide:セレブロシド)の略。
ガングリオシドは、スフィンゴ糖脂質の代謝で、ラクトシルセラミド(Galβ1-4Glcβ1-1Cer)から、合成されるが、ABO式血液型物質や、グロボシドも、ラクトシルセラミドから、合成される。パラグロボシドは、ガラクトース(Gal)と、N-アセチルグルコサミン(GlcNAc)からなる、タイプ1の基幹領域(Galβ1→3GlcNAc)を、含んでいて、フコース(Fuc)が結合すると、O型抗原(H抗原)になる。
セラミド(Ceramide:Cer)は、セレブロシド(cerebroside)とも呼ばれる。グルコセレブロシド(glucocerebroside)は、グルコシルセラミド(glucosylceramide)とも呼ばれる。
Fabry病で蓄積するトリヘキソシルセラミド(trihexosylceramide)は、別名、グロボトリオシルセラミド(globotriosylceramide:Gb3)、グロボトリアオシルセラミド(globotriaosylceramaide)、セラミドトリヘキソシド(ceramide trihexoside:CTH)、とも呼ばれる。
Gb3は、細胞表面のベロ毒素受容体でもある。
注5:グルカン(Glucans)は、ブドウ糖(グルコース)のみが結合した多糖類で、α-グルカンと、β-グルカンとがある。
β-D-グルカンは、肝臓のクッパー細胞(Kupffer細胞)をも、活性化する。
キノコ類(シイタケ、ナメコなど)の中には、β-(1-3)-Dグルカンや、β-(1-6)-Dグルカンが、多く含んでいる。
βグルカンは、真菌などの表面(表層)を形成している。
βグルカンの受容体(レセプター)には、Dectin-1が、ある。
注6:免疫系は、外敵の病原体(細菌など)から、自己の体を守るために、発達したシステム。
免疫系で、細菌などの病原体から、自己の体を守るためには、自己の組織(味方)と、非自己の組織(敵)を、識別する仕組みが、存在する。
原始的な獲得免疫である、NK細胞は、NK阻止レセプターで、標的細胞表面に、自己と同じMHCクラスI分子(α鎖)を認識すると、自己(味方)と認識し、攻撃しない。そして、NK細胞は、標的細胞表面に、自己と同じMHCクラスI分子(α鎖)を認識出来ないと、攻撃する。言わば、戦場では、味方(自己)だという紋章をつけていない兵を、敵と見なすように、NK細胞は、自己であると証明するMHCクラスI分子(α鎖)を、発現していない細胞を、非自己(敵)と、見なす手法だ。つまり、NK細胞は、自己(味方)である証明があるか、監視していると、考えられる。
他方、ヘルパーT細胞の様に、T細胞は、TCR(T細胞受容体)で、単球・マクロファージ表面の、MHC(HLA)と抗原ペプチドの複合体を認識する。これは、どちらかと言えば、自己のMHC(HLA)が変性していないか、認識し、変性していなければ自己(味方)、変性していれば非自己(敵)と、認識する。つまり、T細胞は、自己が変性していないか、監視していると、考えられる。これは、言わば、戦場では、味方(自己)が、裏切っていないか(変性していないか)、監視して、敵の紋章を付けていなければ味方(自己)、敵の紋章を付けていれば敵(非自己)と、見なすような手法だ。
このように、原始的な免疫機構であるNK細胞は、KIRなどにより、標的細胞が、MHCクラスI分子を発現しておらず、自己(味方)である証明を有していないと、非自己(敵)と看做して、攻撃する。他方、より進化した免疫機構であるT細胞(ヘルパーT細胞、キラーT細胞)は、TCR(T細胞受容体)により、標的細胞が、ペプチド抗原などにより、変性したMHCクラスI分子を発現していて、自己と変性していると、非自己(敵)と看做す、と考えられる。つまり、標的細胞のMHCクラスI分子は、NK細胞にとっては、自己(味方)であることの証明書であり、T細胞にとっては、自己(味方)が変性していないか監視する対象であると、考えられる。
原始的な免疫機構であるNK細胞は、体内に、自己(味方)である証明を有していない細胞(MHC分子を有しない細胞)が存在しないか監視し、他方、より進化した免疫機構であるT細胞は、自己(味方)である細胞(MHC分子を有する細胞)が、変性していないか監視している。
NK細胞は、自己としての標識(MHCクラスI分子)を欠落した細胞を、異物(敵)として看做す(非自己認識:missing self recognition)。
NK細胞が、非自己認識を行う際には、抑制化レセプター(抑制受容体)が重要な役割を果たす。
NK細胞は、抑制受容体を発現し、自己の主要組織適合性複合体(MHCクラスI分子)を認識し、MHCクラスI分子を発現している正常細胞をは(自己と看做し)攻撃しないが、MHCクラスI分子を発現していない細胞(癌細胞、ウイルス感染細胞)をは(非自己と看做し)攻撃する。
ヒトのNK細胞には、NK細胞受容体(抑制化レセプター)の代表として、killer inhibitory receptor(KIR)であるCD158が存在する。
ヒトのNK細胞には、活性化レセプターとして、natural cytotoxicity receptors(NCR)と呼ばれるNKp44、NKp46、NKp80、NKG2D(CD314)、2B4(CD244)などが存在する。NK2GDはMICやULBPを認識し、2B4はCD48を認識する。
参考文献
・宮坂信之、他:わかりやすい免疫疾患 日本医師会雑誌 特別号(1) 生涯教育シリーズ−67、2005年.
・谷口克、他:標準免疫学(第2版、医学書院、2004年).
・山本一彦、他:カラー図解 靭帯の正常構造と機能 IV 血液・免疫・内分泌 (日本医事新報社、2002年).
・森本幾夫:Tリンパ球の免疫における役割 日本医師会雑誌 第123巻・第11号、1733-1739, 2000年.
・宮坂信之:ナチュラルキラー細胞の働き 日本医師会雑誌 第123巻・第11号、1740-1742, 2000年.
・岸本忠三、他:岩波講座 免疫科学3 免疫担当細胞(岩波書店、1986年).
・金田一孝、垣生園子、他:新免疫学叢書10 キラー細胞(医学書院、1983年).
・山村雄一、他:免疫学4 細胞性免疫 アレルギー(中山書店、1982年).
・安保徹:NK-LAK細胞の最近の知見 日本臨床免疫学会雑誌 Vol.15 No.6、517-521、1992年.
・矢田純一:E(赤血球)リセプター 免疫学1 免疫担当細胞 リンパ球表面レセプター、220-225, 1981年(中山書店).
・矢田純一:栄養素と免疫機能 小児科 Vol.39 No.9、999-1005、1998年.
・赤羽太郎、青山香喜、柳沢光彦:特集・新しい臨床検査 リンパ球のSubpopulation 小児科診療 第45巻・第6号、39-46、昭和57年(診断と治療社).
・荒瀬尚:NK細胞受容体、Medical Tribune、2007年1月25日号、37頁.