糖鎖
1.糖鎖とは
グルコース(ブドウ糖)、ガラクトース(Gal)、マンノース、N-アセチルグルコサミン(GlcNAc)、N-アセチルガラクトサミン(GalNAc)、フコース、キシロース、シアル酸などの糖は、複雑に連なって、糖鎖を形成する。
細胞表面の糖鎖は、他の細胞(白血球、癌細胞など)、細菌、ウイルス、毒素などが、細胞に接着する際の結合部位(リガンド)となる。
細菌は、表面のレクチンにより、宿主の糖鎖と結合する。
糖鎖は、蛋白や脂質と結合して、糖蛋白質や、糖脂質となり、結合した蛋白質や脂質を安定化させたり、蛋白質のタグ(荷札)として細胞間での情報伝達に、重要な役割を果たしたり、プロテオグリカンとして水分を結合させ組織を保護する。
糖鎖は、蛋白質に結合して、蛋白質のタグ(荷札)の役割を担う。また、糖鎖は、結合した蛋白質や脂質を安定化させる。シアル酸の付いた糖鎖が結合すると、陰性荷電により、血管内皮細胞と反撥し、肝臓などで、分解されにくくなる。
蛋白質や脂質に糖鎖が結合したものは、複合糖質と呼ばれる。複合糖質には、糖蛋白質、糖脂質、プロテオグリカンに分類される。
糖蛋白は、1本の蛋白質に、短い糖鎖(単糖が20個まで)が、1〜数百本の糖鎖が結合している。
糖脂質は、1本の脂質分子に、1本の糖鎖が結合している。
プロテオグリカンは、1本の蛋白質(コア蛋白)に、長い糖鎖(単糖が100〜1万個:グリコサミノグリカン)が結合している。
糖鎖の単糖成分 ヘキソース ガラクトース(Gal)、マンノース(Man)、グルコース(Glc) デオキシヘキソース L-フコース へキソサミン N-アセチルグルコサミン(GlcNAc)、N-アセチルガラクトサミン(GalNAc) シアル酸 N-アセチルノイラミン酸(NeuAc)、N-グリコリルノイラミン酸(NeuGc) ペントース キシロース、L-アラビノース
糖蛋白質の糖鎖部分は、親水性の供与、蛋白分解酵素からの保護作用、レクチンとの結合、ホルモンや微生物の受容体、細胞間相互作用などがある。
2.糖鎖と癌転移
糖鎖には、O型糖鎖(セリン・スレオニン結合型糖鎖:大きい糖鎖)と、N型糖鎖(アスパラギン結合型糖鎖:小さい糖鎖)とが、存在する。O型糖鎖(O結合糖鎖)は、アミノ酸のセリン(Ser)やスレオニン(Thr)に結合していて、比較的、構造は単純。N型糖鎖(N結合型糖鎖)は、アスパラギン残基に結合していて、シグナルの役割を果たしている。
N型糖鎖の分岐を担う、GnT-V(β-1,6-N-アセチルグルコサミン転移酵素-V)の過剰発現は、癌転移を促進させる。
GnT-IIIを導入した癌細胞では、E-カドへリン(細胞を糊付けする糖鎖)が、増加して、細胞間接着が高まる:E-カドへリンは、細胞内に存在するα、β、γ-カテニンと複合体を形成し、裏打ちされる。癌細胞に、GnT-IIIの遺伝子を導入し、癌細胞表面のE-カドへリンに結合したN型糖鎖に、GlcNAcを発現させると、β-カテニンのチロシンリン酸化と細胞膜から細胞質への移行が、抑制され、E-カドへリンによる細胞間接着が高められ、癌の転移が抑制される。
E-カドヘリンは、細胞間接着分子であり、E-カドヘリンの発現が減少した癌細胞は、リンパ節転移し易い。低分化癌は、高分化癌に比して、癌細胞間接着分子に異常がある(癌細胞間接着機能が低下している)ことが多く、転移し易い。スキルス胃癌、乳癌の小葉癌、低分化前立腺癌などは、形態学的に明瞭な細胞間接着を示さない低分化癌であり、30〜50%の症例で、E-カドヘリン遺伝子異常が起こっている。なお、低分化癌では、細胞基質接着分子(細胞の極性を決定している)にも異常があり、癌が転移し易い。
E-カドヘリンは、Ca依存性の接着分子で、癌化に伴って、E-カドヘリンを合成する遺伝子が修飾され(プロモーター領域のDNAが、メチル化される)、E-カドヘリンの発現が低下し、癌の転移が、促進される。
5アザデオキシシチジンは、カドヘリンを復活させる(カドヘリンの発現が増加する):乳腺癌を有するマウスに、DNAメチル化の阻害剤である5アザデオキシシチジンを投与(注射)すると、カドヘリンを合成する遺伝子の修飾が、除去され、カドヘリンが合成され(カドヘリンが復活させられ。癌細胞表面にカドヘリンが発現し)、乳腺癌の肺転移が、抑制された。
3.糖鎖と癌転移
シアル酸が結合した糖鎖を有する糖蛋白質や糖脂質は、シアル酸の陰性荷電により、陰性荷電を有する血管内皮細胞などと、電気的に反撥し、肝臓での分解を逃れる。
EPO(エリスロポエチン)には、糖鎖が結合している。EPOの糖鎖のシアル酸を外すと、in vitro(試験管内)では、EPOの活性は、3倍になる。しかし、糖鎖を完全になくすと、EPOの活性は、なくなる。また、EPOの糖鎖のシアル酸を外すと、in vivo(生体内)では、肝臓でEPOが分解されてしまい、EPOの活性は、なくなる。
遺伝子工学の遺伝子組み換え技術(recombinant DNA technology)で合成した蛋白質は、糖鎖を結合させないと、本来の生理活性を持った蛋白質にはならない。
4.糖鎖と先天性筋ジストロフィー
先天性筋ジストロフィーでは、筋膜が脆弱で、筋肉が収縮を繰り返すと、筋膜が破壊し、筋肉も崩壊して、最終的に、筋萎縮が起こる。
筋肉細胞は、筋膜(筋細胞膜:sarcolemma)の直下(筋細胞内)に、ジストロフィン(dystrophin)と呼ばれる蛋白質が存在し、アクチン(筋原線維としてでなく、細胞骨格としてのアクチン)と結合している。
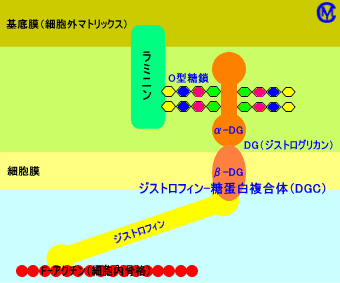
ジストロフィンには、ジストログリカン(DG:dystroglycan)と呼ばれる筋細胞膜の糖蛋白が、結合し、ジストロフィン-糖蛋白複合体(DGC)を形成している。
ジストロフィン-糖蛋白複合体(DGC)を構成しているジストログリカンは、筋肉細胞外(筋線維外)の基底膜のメロシン(ラミニン:laminin α2)に、結合している。
このように、筋細胞の細胞内骨格(アクチン細胞骨格)は、筋細胞外の基底膜(細胞外マトリックス)と、ジストロフィン-糖蛋白複合体(ジストロフィンとジストグリカンの複合体)により、架橋構造を形成している。
架橋構造:細胞外の基底膜−ラミニンα2−O型糖鎖−ジストログリカン−ジストロフィン−アクチン(筋細胞)
このようなジストログリカン(筋膜上に存在する)による架橋構造により、筋細胞の細胞膜(筋膜)は、基底膜(基底層)に、固定されている。
架橋構造では、ジストログリカン(α-DG)は、その表面に存在する糖鎖(O型糖鎖)により、基底膜のメロシン(ラミニン)と結合している。
糖蛋白質(ジストログリカン)の糖鎖の異常は、ある種の先天性筋ジストロフィーの原因になる。
ジストロフィン-糖蛋白複合体(DGC)は、筋細胞の細胞内骨格(アクチン)を、細胞膜(筋細胞膜)を挟んで、筋細胞外の細胞外マトリックス(基底膜)とを結合させ、細胞膜を安定させたり(基底膜側に固定する)、神経筋接合部の構成に関与している。
ジストロフィン-糖蛋白複合体(DGC)に異常があると(ジストログリカンの糖鎖の異常など)、細胞内骨格と、細胞外マトリックスとの結合が低下し、筋肉が、収縮/弛緩した際、筋細胞膜からの細胞内Ca2+流入が増加して、筋細胞が壊死に陥ると言われる。先天的筋ジストロフィーの多くの疾患では、膜(筋細胞膜や、基底膜など)に関連する蛋白に異常があり、筋肉の強い収縮に耐えかねて、膜の破綻が生じる。
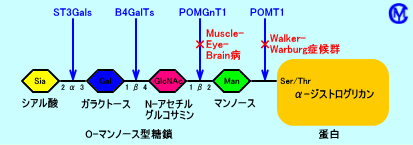
α-ジストログリカンは、蛋白(コア蛋白)に、糖鎖が結合した構造をしている。α-ジストログリカンの蛋白(コア蛋白)に結合した糖鎖は、O型糖鎖のO-マンノース型糖鎖(Siaα2-3Galβ1-4GlcNAcβ1-2Man-O-Ser/Thr)。このO型糖鎖は、短い糖鎖であり、マンノース(Man)、N-アセチルグルコサミン(GlcNAc)、ガラクトース(Gal)、シアル酸(Sia)から構成される。α-ジストログリカンに結合したO型糖鎖は、α-ジストログリカンが基底膜のメロシン(ラミニンα2)と結合するのに、必要。なお、α-ジストログリカンには、N型糖鎖(少ない)や、グルコサミン(GlcN:わずか)も、結合している。
筋細胞膜に存在するα-ジストログリカンは、基底膜(細胞膜を保護する)のラミニン/メロシンと、糖鎖を介して(糖鎖部分で)、結合し、また、細胞内のジストロフィン(細胞骨格のアクチンと結合している)と、結合して、軸を形成する。この軸は、細胞膜を挟んで、細胞外の基底膜と、細胞内の細胞骨格を、結合・固定する。なお、細胞膜にはインテグリン(インテグリンα7β1D)も存在し、基底膜のメロシンと、結合し、細胞外の基底膜と、筋細胞膜とを、結合・固定する。
α-ジストログリカンに結合する糖鎖を合成する糖転移酵素が欠損すると、筋肉のみならず、脳や、眼の病変(異常)を、伴なう。神経系では、神経細胞を固定出来ない(基底膜との固定が脆弱になる)ので、大脳の6層構造が乱れたり、網膜の形成が異常になる。
POMT1(protein-O-mannosyltransferase:O-Man転移酵素)は、蛋白(コア蛋白:α-ジストログリカン)に結合する糖鎖を合成する最初の段階で、マンノース(Man)を、蛋白(コア蛋白)に結合させる酵素。Walker-Warburg症候群(WWS)の一部の症例は、POMT1の欠損が、原因。Walker-Warburg症候群は、先天性筋ジストロフィーの症状に加え、丸石様滑脳症(脳瘤、水頭症、無脳回、脳幹部低形成など)、眼病変(小眼球症、網膜異形成、網膜剥離、視神経低形成、白内障、緑内障など)を、来たす。なお、POMT1の欠損以外にも、少なくとも、フクチン(fukutin)遺伝子、FKRP(fukutin related protein)遺伝子の異常も、Walker-Warburg症候群を来たす。
POMGnT1(UDP-N-acetylglucosamine: protein O-mannose β1,2-N-acetylglucosaminyltransferase:O-linked mannose beta 1,2-N-acetyl-glucosaminyltransferase:GlcNAc転移酵素)は、糖鎖を合成する第2段階で、マンノース(Man)に、N-アセチルグルコサミン(GlcNAc)を結合させる酵素。筋・眼・脳病(muscle-eye-brain病:MEB)は、POMGnT1の欠損が、原因。筋・眼・脳病は、福山型先天性筋ジストロフィー(FCMD)に類似した疾患で、進行性の近視、網膜変性、視神経萎縮が、認められる。
日本に多い、福山型先天性筋ジストロフィー(FCMD)は、FCMDと言う酵素が欠損し、フクチン(fukutin)と呼ばれる蛋白の合成が、行われない。フクチン(fukutin)の機能は、今だ十分に解明されていないが、糖鎖修飾に関係している(フクチンは、細菌の糖転移酵素とアミノ酸配列が似ている)。福山型先天性筋ジストロフィーは、常染色体性劣性遺伝をする。保因者は、通常、発症しない(異常な遺伝子を有する保因者は、正常な遺伝子によりフクチンが生成されるので、フクチン量が半分でも、発症しない)が、例外的に発症する保因者も存在する。福山型先天性筋ジストロフィーの発症には、男女差は、あまりない。FCMDは、骨格筋、心筋、脳(、眼)に、優位に発現している。
フクチン(fukutin)遺伝子は、ヒトの第9染色体の長腕に存在している。
フクチン(fukutin)は、ゴルジ装置(Golgi)に存在している膜蛋白を考えられている。
フクチン(fukutin)は、糖合成酵素の一つで、糖鎖修飾に関与していると考えられている。
フクチン(fukutin)遺伝子は、α-ジストログリカン(α-dystroglycan:α-DG)への糖鎖付加に関与する(作用部位は不明)。
先天性筋ジストロフィーは、ジストログリカン形成に、先天的な異常がある(ジストログリカン異常症)。
Duchenne型筋ジストロフィーは、ジストロフィン形成に、先天的な異常がある。Duchenne型筋ジストロフィーは、乳児期は、遠位の筋肉(下腿の筋肉等)が、運動後などに、腫脹して、肥大(仮性肥大:pseudohypertrophy)する。筋萎縮により、筋力低下が認められない時期でも、血液検査て、CPK、AST(GOT)、ALT(GPT)などの酵素が、著名に上昇している。
ジストログリカン(dystroglycan)自体の欠損が原因の疾患は、存在しない(ジストログリカンが欠損しても、致死的でないと考えられて来た)。
しかし、マウスで、ジストログリカンの遺伝子を破壊した胎児は、致死的で、ジストログリカンは、細胞外マトリックス蛋白質の集合に、必要不可欠だと言う。
5.悪液質とジストロフィン
癌患者では、悪液質(cachexia)になると、骨格筋が萎縮する。
悪液質では、骨格筋のジストロフィン糖タンパク質複合体(dystrophin glycoprotein complex:DGC)が、変化し(ジストロフィンが減少し、DGC蛋白の糖鎖合成が増加する)、筋肉が萎縮すると言う。
6.ヒストン脱アセチル化酵素と筋ジストロフィー
ヒストン脱アセチル化酵素(ヒストンデアセチラーゼ、Histone Deacetylase:HDAC)は、細胞核のヒストンの脱アセチル化を行う酵素。ヒストンが脱アセチル化されると、ヌクレオソーム構造が元に戻り、遺伝子は不活性化される。
筋ジストロフィー(MD)のモデルマウスに、ヒストン脱アセチル化酵素阻害薬であるtrichostatin A(トリコスタチン A)を投与すると、骨格筋量が回復し、筋ジストロフィーに特徴的な筋機能の低下が阻止される(Lorenzo Puri等)。ヒストン脱アセチル化酵素阻害薬を投与すると、筋収縮に関連する筋変性が起こり難くなる(筋変性に対する抵抗性が増大する)。
抗癲癇薬のバルプロ酸ナトリウム(Valproic acid:VPA、商品名:デパケン錠など)は、ヒストン脱アセチル化酵素(HDAC)阻害作用がある。バルプロ酸は、histone deacetylase(HDAC1)を直接的に阻害する(ヒストンのアセチル化が亢進する)。バルプロ酸は、胎児の奇形を誘発する作用(valproic acid-induced birth defects)があるが、このようなバルプロ酸の奇形誘発作用は、バルプロ酸のHDAC阻害作用と関連があると考えられている。
遺伝子の転写に際して、クロマチンの構造が、転写因子と結合出来るように調節される。
クロマチンの構造は、DNAメチル化と、ヒストン修飾(アセチル化、リン酸化、メチル化によるヒストン蛋白修飾)により調節される(エピジェネリック制御)。アセチル化ヒストンに富むクロマチン領域は、転写活性が高いと言われる。
ヒストンのアセチル化は、ヒストンアセチルトランスフェラーゼ(HAT:histone acetyltransferase)により触媒される。ヒストンは、特に、アミノ基末端ドメイン内のリジン残基がアセチル化され易い。
アセチル化されたヒストンは、電荷が中性となる。そして、ヒストン-DAN間や、ヒストン-ヒストン間の結合が緩み、遺伝子が転写されやすくなる。
7.TGFβと筋ジストロフィー
トランフフォーミング増殖因子β(TGF-β)は、TGF-βは、多くの組織で、細胞外基質蛋白を産生し、分解酵素を抑制し、創傷治癒を促進する。
マルファン症候群では、TGF-β(TGFβ)の活性が過剰に発現している。
降圧剤(降圧薬)のロサルタンは、TGF-β活性阻害作用がある。
ロサルタンを投与すると、マルファン症候群発症マウスの大動脈(壁)が強化され、大動脈瘤の発症が抑制される。
筋ジストロフィー(DMD)発症マウスに、TGF-β阻害薬やロサルタンを投与すると、筋肉の損傷を再生する能力が回復すると言う。
おまけ
筋萎縮性側索硬化症(ALS)は、ビタミンE、あるいは、多価不飽和脂肪酸(PUFA)を、毎日、多く摂取すると、発症リスクが、約50〜60%低減すると言う。フラボノイド、リコピン、ビタミンB、ビタミンC、グルタミン酸、カルシウム、植物性エストロゲンは、ALS発症に、関連はないと言う(Journal of Neurology, Neurosurgery and Psychiatry, 2006年:オンライン版)。
参考文献
・吉岡三恵子:総説 先天性筋ジストロフィー研究の新しい展開 日本小児科学会雑誌 109巻6号、703-713, 2005年.
・松尾雅文:ジストロフィン研究の新展開 日本小児科学会雑誌 97巻3号、655-658, 1993年.
・落合淳志:癌の組織学的分化度と接着分子(質疑応答) 日本醫事新報 No.4152(2003年11月22日)、93-94頁.
・清水輝夫:筋ジストロフィーと糖鎖 日本醫事新報 No.4214(2005年1月29日)、1-9頁.
・斎藤加代子、他:ジストロフィンと筋ジストロフィー 小児内科 Vol.23 No.11、1791-1798、1991年.
・斉藤義朗:総説 福山型先天性筋ジストロフィー研究の最近の進歩 日本小児科学会雑誌 110巻3号、389-397、2006年.
・ビタミンEとPUFAで筋萎縮性側索硬化症を予防、Medical Tribune、6頁、2006年7月27日号.
・筋ジストロフィーの筋変性を阻止 ヒストンデアセチラーゼ阻害薬投与マウスで、Medical Tribune、Vol.40 No.1、1頁(2007年1月4日号).
・斎藤博久:第109回日本小児科学会学術集会 分野別シンポジウム:乳幼児気管支喘息治療の早期介入 アレルギー性炎症性疾患に関わる遺伝的背景、エピジェネティック制御と環境因子、日本小児科学会雑誌、111巻1号、10-15頁(2007年).
・降圧薬に筋ジストロフィー治療の可能性、Medical Tribune、2007年8月9日、42頁.
・Acharyya S, et al: Dystrophin glycoprotein complex dysfunction: a regulatory link between muscular dystrophy and cancer cachexia. Cancer Cell. 2005 Nov;8(5):421-32.
・Karen A. Lapidos, et al: The Dystrophin Glycoprotein Complex Signaling Strength and Integrity for the Sarcolemma. Circulation Research. 2004;94:1023.