�@
�@ �@
�@ �@
�@ �@�@�@
�@�A�f��H
�@���Ǔ��ȂǂŎY�����ꂽ�A�����j�A�́A���t�����̑��ɗA������A�A�f�A�O���^�~���_�A�O���^�~���ɕϊ������B
�@�A�f��H�́A�̑��̊̍זE�ɑ��݂��A�̓��Ő������ꂽ�L�łȃA�����j�A���A���łȔA�f�ɕϊ�����B
�@���̂́A�_�o�Ő�������L�łȃA�����j�A���A�Ő��̂Ȃ��O���^�~���Ƃ��āA������A������B
�@�P�D�A�f��H
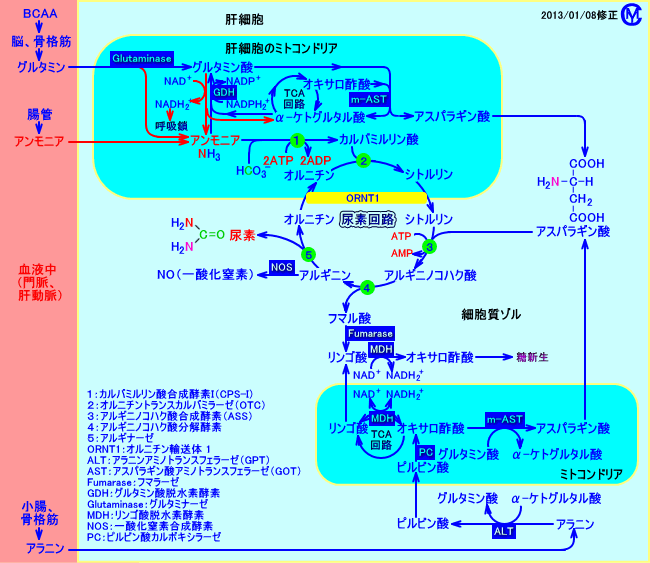
�@�A�f��H�́A�I���j�`����H�A�A�f�T�C�N���A�I���j�`���T�C�N���AKrebs-Henseleit cycle�A�Ƃ��Ă��B
�@�A�f�̍����́A�̑��i�喬���͂̊̍זE�j�̔A�f��H�ŁA�s����B
�@�A�����j�A�Əd�Y�_�C�I������A�J���o�~�������_�����y�f�iCPS-I�A���P�j�ɂ��A�J���o�~�������_�i�J���o���C�������_�j���A���������B
�@�J���o�~�������_���A�I���j�`���iOrnithine�j�ƁA�I���j�`���g�����X�J���o�~���[�[�iOTC�A���Q�j�ɂ��A�������A�V�g������(Citrulline�j�������ߒ��́A�~�g�R���h���A���ōs����B����ȊO�̔����́A�~�g�R���h���A�O�̍זE���]���ōs����
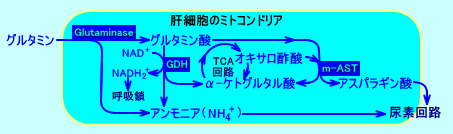
�@�A�f��H�ɕK�v�ȃA�X�p���M���_�́A�A���j�����A�~�g�R���h���A�O�i�זE���]���j�ŁAALT�ɂ��A��-�P�g�O���^���_�ɃA�~�m��]�ڂ��A�������ꂽ�O���^�~���_���A�~�g�R���h���A���ŁAm-AST�im-GOT�j�ɂ��A�I�L�T���|�_�ɃA�~�m��]�ڂ��A���������i���R�j�B
�@�A���M�j�������������ۂɁA�A���M�j�m�R�n�N�_����V������t�}���_�́A�זE���]���icytosol�j�ŁA�t�}���[�[�ɂ�胊���S�_�ɕϊ�����A����ɁA�I�L�T���|�_�ɕϊ�����A���V���ɗ��p�����i�t�}���_���A�~�g�R���h���A���ɗA������āA�A�X�p���M���_���Đ������̂ł͂Ȃ��j�B
�@�A���M�j������́ANOS�iNO synthase�F��_�����f�����y�f�j�ɂ��A�����_�f�i�t���[���W�J���j�̈��ł���ANO�i��_�����f�j���Y�������B
�@�A�f��H�ł́A�I���j�`�����P��]���āA�P���q�̔A�f�����������ɂ́A3ATP��������F�A�����j�A���J���o�~�������_�ɂȂ�o�H��2ATP��2ADP�ɏ����A�V�g���������A���M�j�m�R�n�N�_�ɂȂ�o�H��1ATP��1ADP�ɏ�����F�v4�̍��G�l���M�[�����_������������i���S�j�B
�@���l�ł́A1��30g�̔A�f���A�r�������i120g�̒`�������ى������j�B���̔A�f������ׂɁA�̑��ł́A2mol��ATP�i�̑���1���̃G�l���M�[��ӂ̖�15���ɑ�������j��������B��b��ӗ��i85mol��ATP�j�̖�2�����A�A�f��H�ŏ�����B
�@�R����H�iCori cycle�j�ŁA���_2���q����A�O���R�[�X1���q������ɂ́A�s���r���_����̓��V���ׂ̈ɁAATP6���q���K�v�F�O���R�[�X�����_�ɂ܂ŕ��������ۂɂ́AATP2���q������������Ȃ��̂ŁA�K�v��ATP�́A�~�g�R���h���A���ċz���Ő��������B
�@�O���R�[�X-�A���j����H�ŁA�A���j��1���q���炩��A�O���R�[�X1���q������ɂ́A�s���r���_��������V���ׂ̈�ATP6���q���K�v�ŁA���A�A���j��������o�����A�~�m���f��A�f��H�ŔA�f�Ƃ��ď�������ׂɁAATP2���q���K�v�ŁA���v�AATP8���q��������B
�@�Ȃ��A�t���́A�V�g�������ȍ~�̔A�f��H�̌o�H�����̂ŁA�t���ł��A�����̃V�g����������A�A�f�����������F�t���́A�~�g�R���h���A�O�i�זE���j�̔A�f��H�œ����A�R�̍y�f�i��}�̂R�A�S�A�T�̍y�f�j��L���Ă���̂ŁA�����̃V�g���������A�A�f�ɕϊ��o����B�������ł��A�V�g���������A�O���^�~���_�ƃA�����j�A�Əd�Y�_�C�I�����琶������A�z���t���ɕ��o����A�t���ŁA�A���M�j�������������i���T�j�B �@�Q�D�̑��ŏ��������A�����j�A
�@�Q�D�̑��ŏ��������A�����j�A
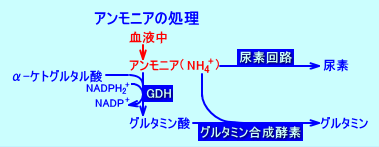
�@�̑��́A�z�����i�喬�����j���A�A�����j�A���O���^�~�����A��荞�ށB
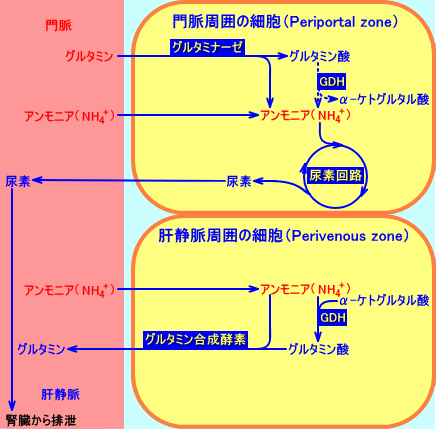
�@�喬���͂̍זE�iperiportal cells�j���ŁA�O���^�~���́A�O���^�~�i�[�[�ɂ�蕪������A���̌��ʐ��������A�����j�A�́A�z�������̑��ɗ���ė����A�����j�A�Ƌ��ɁA�A�f��H�ŁA�A�f�ɕϊ������B
�@�z�������̑��ɗ���ė����A�����j�A�́A�喬���͂̍זE�iperiportal cells�j���ŏ�������Ȃ��ƁA�̐Ö����͂̍זE�iperivenous cells�j�ŁA�O���^�~�������y�f�ɂ��A�O���^�~���ɕϊ������B
�@�]���āA�̑��́A�A�����j�A����荞��ŁA�A�f��r�����邪�A�i�喬�����Ɗ̐Ö������ƂŁj�O���^�~���̔Z�x�́A�قƂ�Ǖω����Ȃ��B
�@�P�j�D���Ǔ��ł̃A�����j�A�Y��
�@���Ǔ��ł��A�A�����j�A���Y�������F���Ǔ��ł́A�H���i�H�a�A�H���j�R���̃A�~�m�_���A�S���O���^�~�i�[�[��GDH�i�����j�A�ۃf�A�~�i�[�[�i�咰�j�ɂ��A��������A�܂��A�A�f���ۃE���A�[�[�i�咰�j�ɂ�蕪������A�A�����j�A���A���������B
�@�����A�����j�A�̑啔���́A���ǁi�����ǁj���ŁA�����S����咰���ۂɂ��Y�����ꂽ�A�����j�A�ƌ������B�����S���O���^�~�i�[�[�ɂ�萶�������A�����j�A�́A���Ǔ��ŎY�������A�������j�A�ʂ̖�1/2���߂�B
�@���Ǔ��ŎY�����ꂽ�A�����j�A�́A�z������āi���U�j�A�喬�������̑��ɗA������A�A�f��H�ŔA�f�ɕϊ����ꂽ��A�O���^�~���_��O���^�~�������������B
�@�Ȃ��A�喬-�̏z�Z���H������A�̍d���ł́A���̌o�H�����B���āA�喬�����A�̏z�ɓ���A���A�����j�A���ǂ�������B
�@�Q�j�D�A�~�m�_�̈ى��ɂ��A�����j�A�Y��
�@�̓��ł́A�`������ӂ̉ߒ��ŁA�A�~�m�_�̈ى��i�E�A�~�m�����j�ɔ��Ȃ����f��ӂŁA�A�����j�A�����������B
�@�Ȃ��A�]�⍜�i�ł́A�O���^�~�������y�f�Ȃǂɂ��A�A�����j�A�͏�������A�O���^�~����A���j���i���V�j�Ƃ��āA�����ɕ��o����A�t���A�����A�̑��ő�ӂ����B
�@�R�j�DGDH�ɂ��A�����j�A�Y��
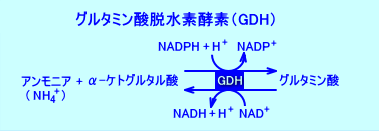
�@�̑��̃~�g�R���h���A���ŁAGDH�i�O���^�~���_�E���f�y�f�j�́A�A�����j�A���������A�O���^�~���_������F�A�����j�A�́AGDH�̕��t���O���^�~���_�iGlu�j�����̕����ɂ���B�A�����j�A�́AGDH�ɂ��O���^�~���_�����i�A�~�m�_���������j�ɕK�v�Ȃ̂ŁA�i�̑����́j�A�����j�A�Z�x���A�ɒ[�ɒቺ���邱�Ƃ́A�̓��ō���������K�{�A�~�m�_�̔Z�x���A�ቺ�����Ă��܂������ꂪ�����B���̂悤�ɂ��āAGDH�́A�A�����j�A����������B�Ȃ��AGDH�́A�t���i�A�ǍזE�j�A�]�A���ɂ����݂��Ă���B
�@GDH�̔����͉t�I�ŁA�A�����j�A�ƃ�-�P�g�O���^���_����A�O���^�~���_������̂Ƃ͋t�ɁA�O���^�~���_���ى��initrogen liberation�j���āA�A�����j�A�ƃ�-�P�g�O���^���_�Ƃɕ�������ى��������AGDH�ɂ��N���Ȃ���i�^�����Ȃǁj�B���̂悤�ɂ��āAGDH�́A�A�����j�A���Y���i�����j����.�B�������AGDH�̕��t�萔�́A�O���^�~���_�������鑤�ɕ��Ă���B
�@�]�ȂǂŐ������ꂽ�O���^�~���́A�̑��ŁA�O���^�~�i�[�[�ɂ��A�A�����j�A�ƃO���^�~���_�ƂɁA��������A�A�����j�A���Y���i�����j�����i���W�j�B�O���^�~���_�́A����ɁAGDH�ɂ�蕪������A�A�����j�A���Y�����ꂽ��Am-AST�ɂ��A�A�f��H�Ŏg�p�����A�X�p���M���_�ɕϊ������B
�@�R�D�A�f��H�ȊO�̃A�����j�A����
�@���t���̃A�����j�A�́A�̑��ŁA��ɁA�A�f��H�ŔA�f�����A���������i�A�����j�A�̏����\�͂́A�A�f��H���A�ł������j�B
�@�A�����j�A�́A�A�f��H�ŏ��������ȊO�ɁA�̑��ŁAGDH�ɂ��O���^�~���_��������A�O���^�~���_�����y�f�iglutamine synthetase�j�ɂ��O���^�~�������A���������B
�@��V�I��OTC�i�I���j�`���g�����X�J���o�~���[�[�j����������AOTC�������ł́A�A�f�������i�܂Ȃ��ׁA�����A�����j�A�l���㏸����i2,000��mol/L�ȏ�j�B
�@OTC�����ǂ̈�`�`���́AX�A���i������`�j�����A�����i�w�e���ڍ��́j�����ǂ���̂ŁA�j����́A�P�F�P�Ȃ����A�P�F�Q�Ƃ����iX-linked semidominant�j�B
�@OTC�����ǂ̔��Ǘ��́A�č��łP�F�T�O�C�O�O�O�A���{�ł͂P�F�W�O�C�O�O�O�Ƃ����B
�@OTC�����ǂ́A�V���������ɔ��ǂ��āA����P�T�Ԉȓ��Ɏ��S���邱�Ƃ�����B
�@OTC�����ǂ́A���M�A�Q��A��J�Ȃǂ̗v���ŁA����i���A�����j�A���ǁj�������N�����A���C�l�nj�Q�̏Ǐ��悵�Ď��S���邱�Ƃ�����B
�@�j�����҂̕�e�́A�ۈ��҂̂��Ƃ������A�������҂̕�e�́A�قƂ�Ǖۈ��҂łȂ��B
�@���҂̔����͐e�i�قƂ�ǂ���e�j����̈�`�A�c��̔����͐V�����ˑR�ψق������ƌ�����B
�@OTC��`�q�̓ˑR�ψق́A��e�����A���e�̐��B�זE�ŋN����₷���B
�@OTC�����ǂ̊��҂́A���A�����j�A���ǂ̂��߁A���ӊ��A�q�f�Ȃǂ̏Ǐ�Ŕ��a���A����ɁA�����ُ�A�����A���͏�Q�A�ӎ���Q��悷��B���t�A�Ìňُ�ȂǁA�̍זE�̎�����Q�̏����́A�����łȂ��B
�@��V���A�f��H�ُ�ǁi��V���A�f�T�C�N���ُ�ǁj�̓��AOTC�����ǁi�I���j�`���g�����X�J���o�~���[�[�����ǁj�ƁACPS�����ǁi�J���o�~�������_�����y�f�����ǁj�ł́A���A�����j�A���ǂ�������ۂɁA�������J���j�`�����ቺ���A�A�V���J���j�`��/���J���j�`���䂪�㏸����i�J���j�`�����R�́A���A�����j�A���ǂ�������j�B �@�T�D�O���^�~���_�́A���f�̈ى��ɂ��A�����ɂ��d�v�Ȓ��ԑ�
�@�T�D�O���^�~���_�́A���f�̈ى��ɂ��A�����ɂ��d�v�Ȓ��ԑ�
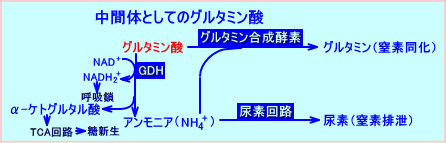
�@�O���^�~���_�́A�O���^�~�����������ăA�����j�A����������ہi�A�~�m�_�����j�ɂ��A�A�����j�A��A�f��H��������ہi�A�~�m�_�ى��j�ɂ��A���ԑ̂Ƃ��āA�֗^����B
�@�O���^�~���_�́A���f�r���̌o�H�ł́A���f���������initrogen donor�j�A�̑���GDH�Ŏ_���I�E�A�~�m������A�A�����j�A������B�t�ɁA�O���^�~���_�́A���f�����̌o�H�ł́A���f����������initrogen acceptor�j�A�O���^�~�������y�f�iglutamine synthase�j�ɂ��A�O���^�~���ɕϊ��iconvert�j�����B�O���^�~���́A�t���̔A�ǍזE�ɗA������A�O���^�~�i�[�[��GDH�iglutamate dehydrogenase�j�ɂ��E�A�~�m�������B���́A�t���ŁA�O���^�~�����A�O���^�~�i�[�[��GDH�ɂ��E�A�~�m�������ۂɁA�A�����j�A�������i�Y���j����A�A�����j�E���C�I���iNH4+�j�Ƃ��ĔA���ɔr�������B���̃A�����j�E���C�I���̔r���́A�A���A���J�������A�A��pH�𐳏�ipH4�`pH8�j�Ɉێ�����B
�@�����g�D��A�̑���GDH�ɂ��A�A�����j�A�����ʂɎY�������ƁA�_�o�Ő���A�A���J���[�V�X���A�����B
�@GDH�ƃO���^�~�������y�f�iglutamine synthatase�j�́A���Ă̔������iorganisms�j�ɑ��݂��A�A�����j�A���O���^�~���_��O���^�~���ɕϊ�����̂ɗL�p�B
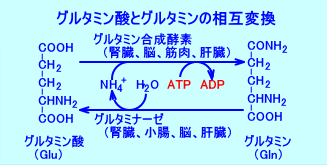
�@�Ȃ��A�O���^�~���_�ƃO���^�~���́A�O���^�~�������y�f�ƁA�O���^�~�i�[�[�ɂ��A���݂ɕϊ������B
�@�U�DBUN�i�A�f���f�j
�@�̑��̔A�f��H�Ő������ꂽ�A�f�́A���t����A������A�t���̎����̂��h�߂���A�A���֔r�������B
�@�A�f�́A�̊O�ɔr������钂�f�ʂ�80�`90�����߂�B
�@���t�i�����j���̔A�f�̒��f�ʂ́A�A�f���f�iBUN�FBlood Ures Nitrogen�j�Ƃ��āA���������B
�@BUN�́A�t�@�\��Q�i�t�s�S�j������Ə㏸����i�t������̔r���̏�Q�j�BBUN�́A�܂��A��H�i�g�D�`���̈ى��̘��i�j�A���M�A�E���i�������^���A�����A�q�f�Ȃǁj�A���`���H�ł��A�㏸����B
�@BUN�́A�̕s�S�A�D�P�A��`���H�ł́A�ቺ����B
�@BUN�́A�������̔Z�x���A�S�����̔Z�x�̕����A���Ⴂ�B
�@BUN�́A���f�ʂƂ��đ��肵���l�ɁA2.14�i��60/28�j���悶���l���A�A�f���f�iBUN�j�̑���l�Ƃ��āA�������B�@
�@NPN�iNon-Protein Nitrogen�j�Ƃ́A��`�������f�������̑����f�ʂ̂��ƁB
�@NPN�́A�������̔A�f�i18�`42mg/dl�j�A�A�_�i2�`7mg/dl�j�A�N���A�`���A�N���A�`�j���A�V���A�~�m�_�A�A�����j�A�Ɋ܂܂�钂�f�̍��v�i���ʁj�BNPN�̖����ABUN�i�A�f���f�j�̒��f�B�N���A�`���́A�A�~�m�_�������ŁA�̑���t���ōŏI�I�ɐ�������A�ؓ��Ɏ�荞�܂��B�ؓ��̃N���A�`���̖�Q�����x�́A�����q�����āA�s�t�I���N���A�`�j���iCre�j�ɕω�����B�ؓ�������o���ꂽ�N���A�`�j���́A�t������A�A���ɔr�������B�N���A�`�j���́A�N���A�`���ƈقȂ�A�t���̔A�ǂ���A�ċz������Ȃ��B���̂悤�ȃN���A�`�j���̐����𗘗p���A�����N���A�`�j���l�iCre�j�́A�t�@�\��Q�̎w�W�Ƃ��āA�p������B�`����ӂŐ��������A�f���f�l�iBUN�l�j���A�t�@�\�̎w�W�ɂȂ邪�A�N���A�`�j���l�́A�H���ɂ��`���ێ�ȂNJO�I���q�̉e�������܂�Ȃ��ƌ����Ă���B
�@�V�D���̑�
�@�E�M���ނł́A�_�o�Ő�������A�����j�A�i�A���J�����j���A���Q�ȔA�f�i�����j�ɕς��āA�A���ɁA�r������B
�@���f��ӎY���́A�O�������ł̓A�����j�A�Ƃ��āA�C�������ނ�W�������ނ◼���ނł͔A�f�Ƃ��āA��ނⒹ�ނł͔A�_�Ƃ��āA�r�������B
�@���f��ӎY���́A�M���ނł́A�A�_�Ƃ��Ă��A�ĂсA�A�f�Ƃ��āA�r�������B����́A�M���ނ��A�A�Z�k�@�\�Ƃ��āA�A�f�𗘗p���A��Z�k����@�\���\�z���Ă��邱�ƂƁA�֘A������B
�@�E�A�f�́A���q�ʂ�60�Ə������A�זE���O�����R�Ɋg�U����B
�@�A�f���A�a�C�ŁA�̓��ɒ~�ς���ƁA�̓����Z�������A�e������B
�@�����Z�����́A��������Na�Z�x�A�����l�ABUN�l����A���L�̌����Z�����̊T�Z����p���āA�T�Z�o����B�@
�@�����Z�����iOsm/L�j������Na�imEq/L�j�~1.86�{�����l�img/dL�j��18�{BUN�img/dL�j��2.8
�@���t���̔A�f���f�iBUN�j��2.5mg/dL��������ƁA�Z������1mOsm/L��������B
�@�E�A�f�́A�����Ȃ̂ŁA�_����t�ɂ́A�قƂ�NJ֗^���Ȃ��B
�@�@
�@�E�A�f�����t���ɑ�������ƁA�����̋@�\���}������A���t�́A�Ìł���Ȃ�B
�@�E�A�f�́A���n���łȂ��ׁA�זE���i������d�w�j��ʉ߂��ɂ����̂ŁA�A�f�A���̂ɂ��A�זE����ʉ߂���B
�@�M���ނł́A�A�f�A���̂��A�A�Z�k�@�\�ŁA�d�v�ȓ������ʂ����Ă���B
�@�A�f�A���̂ɂ́AUT-A�ƁAUT-B�Ƃ��A���݂���BUT-B�́AKidd�����t�^�R�������B
�@�E�̓��ŁA�`�����������s���ׂɂ́A�c��ȗʂ�ATP�i�G�l���M�[�j�������B
�@�q�g�̑̂ł́A�����A���`�����̖�2������������i�̏d65kg�̐l�̏ꍇ�A��200g�̒`���������������j�@�A�����Ő�����A�~�m�_�̖�2/3�͒`���������ɍė��p����A�c��1/3�́A���V���⎉�b�_�����ɗ��p���ꂽ��A�G�l���M�[���ƂȂ�i�R�Ă����j�B
�@200g�̒`��������������ׂɂ́A17mol��ATP��������B1���̑���b��ӗ��́AATP�ɃG�l���M�[�ϊ�����ƁA85mol�ɂȂ�B�]���āA200g�̒`��������������ׂɏ�����17mol��ATP�́A��b��ӗʂ�20���ɑ�������B
�@�E���̓��̗]��Ȓ��f�́A�q�g�ł́A�A������A�f�Ƃ��Ĕr�������B
�@���f�����́A���֒��⊾���ɂ��r������邪�A����ȏꍇ�������A�r���ʂ͂Pg/�����x�Ə��Ȃ��B
�@�]���āA�A�������f�r���ʂ́A���̂̑����f�r���ʂƊŘ��B
�@�܂��́A���f�r���ʁig/���j���A���A�f���f�r���ʁig/���j�~1.25
�@�E�n���ɂ́A���q�f�iN2�j���i�Œ�j����\�͂�L����ہi���f�Œ�ہj���A���炵�Ă���B
�@
�@���f�Œ�ہiClostridium pasteurianum�j�́A���f�Ɛ��f�����A�����j�A�iNH3�j����������y�f�j�g���Q�i�[�[��L���Ă���B
�@N2�{3H2��2NH3�@�@�@��H���|21.8kcal�i�|91.2kJ�j
�@�j�g���Q�i�[�[�́A�A�]�t�F���h�L�V���ƃ����u�h�t�F���h�L�V����2��ނ̒`������\������Ă��āA�G�l���M�[���Ƃ���ATP�A���f���Ƃ��ĊҌ��^�t�F���h�L�V����K�v�Ƃ���B
�@�j�g���Q�i�[�[�́A�_�f���������A�_�f�ɂ�芈�����s�t�I�ɑj�Q�����̂ŁA���f�Œ�ۂ́A�ċz�n��`�Ԃ�ω������A�_�f�ɂ��j�g���Q�i�[�[�̊����̎�����h���ł���B
�@�j�g���Q�i�[�[�́AADP�ɂ���Ă���������i�s���������j�B
�@���f���q�iN2�j�́A�O�d������L���Ă��āA���w�I�Ɉ���ȕ��q�Ȃ̂ŁA�������ɂ͍��G�l���M�[��v����B
�@���f�Œ�́A���j�����i�ΐ����C���ہA�D�C���ہA�������ہj�����ōs����B
�@�j�g���Q�i�[�[�̍y�f�����́A�_�f�ɂ��s�t�I�ɕs�����������i�_�f�ɑ��銴�������j�B�]���āA���f�Œ�́A�ΐ����C�������ł���B
�@���C���ۂ�������ۂ̓j�g���Q�i�[�[�n���_�f�ɐG��Ȃ��悤�ɕێ��o���邪�A�D�C���ۂ�ΐF�ۂ͕ێ��ɖ�肪������B
�@�D�C���ہiAzotobacter vinelandii��Mycobacterium flavum�j�́A�_�f�������C�����Ⴍ����ƁA���f�Œ�\���㏸����BAzotobacter ���ł́A�ċz���߁i�ċz�n�ω��j�╪�q�\���ω��i�`�ԕω��j�ɂ��A�j�g���Q�i�[�[���_�f�ɂ��s��������ی삵�Ă���B
�@�����ہi��ᎃo�N�e���ARhizobium�j���A���ȐA���̍��тɊ�������ƁA����4�{�̍זE�����A�����i��ᎁj���`�������B
�@�����ۂ́A�����̒��ő��B�E�c�����A�s�K���Ȍ`��̃o�N�e���C�h�ibacteroid�j�ɕό`����B
�@�o�N�e���C�h�́A�y�뒆�̒��f���A�����j�A�ɊҌ����A�A�����j�A�͐A���זE�����̃O���^�~�������y�f�ɂ��O���^�~���_�Ɣ������O���^�~������������A�O���^�~���̃A�~�h��A�X�p���M���_�ɓ]�ڂ���ăA�X�p���M������������A�A�X�p���M���͍��̗A���n�ɂ�荪��������A�����ɗA�������B
�@���܂��F�A�~�m�_�̒��f�ى��i���f��Ӂj
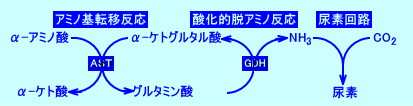
�@�A�~�m�_�̒��f�́AAST�Ȃǂɂ��A�~�m��]�ڔ����itarnsamination�j�AGDH�Ȃǂɂ��_���I�E�A�~�m�����itransamidation�j���o�āA�A�����j�A�ƂȂ�A�A�f��H�ŁA�A�f�ɕϊ����A�r�������B�A�~�m��]�ڔ����ł́A�A�~�m�_�̃A�~�m��iNH2�j���A�P�g�_�i�s���r���_�A�I�L�U���|�_�A��-�P�g�O���^���_�j�ɓ]�ڂ����B
�@�A�~�m��]�ڔ����́AAST�iGOT�j��AALT�iGPT�j�ɂ��s����BALT�iGPT�j�́A���i�ŁA�A���j�������A�̑��ɁA�s���r���_�ƃA�����j�A����������̂ɁA�d�v�Ȗ��������Ă���B�̑��ł́AALT�iGPT�j�́A�A���j���̃A�����j�A����-�P�g�O���^���_�ɓ]�ڂ��A�s���r���_���Đ��iregenerates�j���A�O���^�~���_���A��������B�s���r���_�́A���V���Ɏg�p�����iglucose-alanine cycle�j�B
�@���܂��Q�F�̍זE���̍y�f�̈Ⴂ
�@�̑��ɓ��錌�t�́A�喬����80�����A�̓�������20�����A���������B���̖喬�Ɗ̓����̌��t�́A�ޓ��isinusoid�j��ʉߒ��ɁA�}���ɍ�����A���S�Ö����o�āA�̐Ö��ɗ��o����B�喬���͂̊̍זE�i�喬�ɋ߂����݂���זE�Fperiportal cells�j�́A���V����A�f��H�̍y�f�����������B�����A�ޓ��̊̐Ö����͂̊̍זE�iperivenous cells�j�́A�̍y�f�����������B�A�f��H�ɃA�����j�A������������A���V���𑣐i������O���^�~�i�[�[�́A�����ς�A�喬���͂̊̍זE�ɑ��݂���B�O���^�~�������y�f�́A�̐Ö����͂̊̍זE�ɂ̂ݑ��݂���B�O���^�~�i�[�[�̑��݂���̈�́A�O���^�~�������y�f�̑��݂���̈���A�L���B
�@���܂��R�F�A�X�p���M���_�ƃA�X�p���M��
�@�A�X�p���M���́A�A�X�p���M�������y�f�iasparagine synthase�j�ɂ��A�A�X�p���M���_�ƃO���^�~���̔����ŁA�A�~�m��]�ڂ����B
�@�A�X�p���M���_�{�O���^�~���{ATP���O���^�~���_�{�A�X�p���M���{AMP�{�����_�iPPi�j
�@�A�X�p���M���́A�A�X�p���M�i�[�[�ɂ��A�A�X�p���M���_�ƃA�����j�A�ƂɁA���������B
�@�A�X�p���M���{���iH2O�j�̃A�X�p���M���_�{�A�����j�A�iNH4+�j
�@����ȍזE�́A�A�X�p���M���������o���邪�A�}�������p�������a�זE�́A�A�X�p���M���������o���Ȃ��̂ŁA���B�ɁA�A�X�p���M����K�v�Ƃ���BL-�A�X�p���M�i�[�[�i���i���F���C�i�[�[�j�́A�A�X�p���M�������āA�����a�זE�̑��B���A�}������B
�@���܂��S�F�A�f�̍ė��p
�@�̑��Ő������ꂽ�A�f�̂قƂ�ǑS�ẮA���t��t���ɗA������A�t�����̂��h�߂���A�A���ɔr�������B
�@�A�f�́A�̓��ŁA�ĂсA��ӂɍė��p����邱�Ƃ͂Ȃ��ƍl�����Ă������A�ɔ��ʂ̔A�f�́A�ė��p�����ƌ����B
�@�A�f���o���ێ悳����ƁA�����ŋz������A�Ăя����ɕ��傳��邩�A���邢�́A�����ŋz�����ꂸ�ɑ咰�ɑ����A�咰���������̃E���A�[�[�ɂ�蕪������A�A�����j�A�����������B���̃A�����j�A�́A�咰�ŋz������A�̑��ŁA�A�~�m�_�i�O���^�~���_�Ȃǁj�����������ƌ����B
�@���̂悤�ȔA�f�̍ė��p�́A���ؐH��A���H�����Ă���l�Ō���ꂽ���A����t�H�������Ă���l�ł͌����Ȃ������ƌ����B
�@���܂��T�F���ƐZ����
�@�T���i�L�j��G�C�́A���Z�����̊C�����ŒE������Ȃ��悤�ɁA���t���ɔA�f��~�ς��A�����i�̉t�j�̐Z�������C�����݂ɍ��߂Ă���B�T���̌����A�f�Z�x�́A350mmol/L�i��2g/dL�j�ȏ゠��ƌ�����i�q�g�̌����A�f�Z�x��100�{�j�B
�@�T���́A���G�ȍ\���������t���ŁA�قڑS�Ă̔A�f������i�ċz���j���A�����A�f�Z�x�����߂�B
�@�A�f�́A�J�I�g���s�b�N�C�I���Ƃ��ē����A�זE���̑����̍y�f������j�Q����ƌ�����B�q�g�ł́A�����A�f�Z�x���㏸����ƁA�_�o�זE�Ȃǂ̍זE�̋@�\����Q�����i�A�ŏǁj�B�������A�T���́A�����A�f�Z�x���㏸���Ă��A�זE�̋@�\����Q����Ȃ��B
�@�T�����ɂ́A�A�f��TMAO���܂܂�Ă��āA���M����ƃA�����j�A�L������B�܂��A�T�����́A�Â��Ȃ�ƁA�ۂɂ��A�f���A�����j�A�ɕ��������B
�@�d�����ނ́A�q�g�Ɠ��l�̌����Z�����ȈׁA�C�����ł��ҁi�G���j����E�������ׁA����ɊC��������ł��邪�A�C���Ƌ��ɋz�����ꂽ�����́A�҂ɑ��݂������ލזE����A�C���ȏ�ɔZ�k����āA�r�������B
�@���P�F�J���o�~�������_�����y�f�h�i�J���o���C�������_�V���^�[�[�h�Fcarbamoyl-phosphate synthetase I�FCPS-I�j�́A�A�����j�A�A�d�Y�_�C�I���AATP���k�������A�J���o�~�������_�icarbamyl phosphate�F�J���o���C�������_�j���������锽�����A�G�}����y�f�B
�@�J���o�~�������_�����y�f�h�iCPS-I�j�́AN-�A�Z�`���O���^�~���_�iN-acetyl-glutamate�j�ɂ��A�A���X�e���b�N�ɁA�y�f�������A���i�����i�J���o�~�������_�̍��������i�����j�B
�@���Q�F�I���j�`���g�����X�J���o�~���[�[�i0rnithine transcarbamylase�j�́A�I���j�`���g�����X�J���o���C���[�[�iornithine transcarbamoylase�j�Ƃ��Ă�AOTC�Ɨ������B
�@�I���j�`���g�����X�J���o�~���[�[�iOTC�j�́A�I���j�`���J���o�~���]�ڍy�f�i�I���j�`���J���o���C���g�����X�t�F���[�[�Fornithine carbamoyltransferase�j�Ƃ��A�Ă�AOCT�Ɨ�����邱�Ƃ�����B
�@�A�f��H��OTC�́A�喬���́iperiportal cells�j�̊̍זE�ɑ��݂���B
�@���R�F�̑��^�O���^�~�i�[�[�ɂ��A�O���^�~�����琶�������O���^�~���_���Am-AST�ɂ��A�~�m��]�ڂ��āA�A�X�p���M���_����������A�A�f��H�ŗ��p�����Ƃ����������B
�@�̑��ł́A�A���j����O���^�~���Ȃǂ̒��f���A�A�f��H�ŁA�r������B��V�I�ɔA�f��H�Ɉُ킪����ƁA�������̃A���j����O���^�~���ȂǁA���K�{�A�~�m�_�Z�x���A�㏸����B
�@�Ȃ��A���퐬�l�ł́A�������̃A���j���Z�x�i66mM/dL�j�́A�O���^�~���_�ƃO���^�~���Z�x�i19mM/dL�j���A�����B
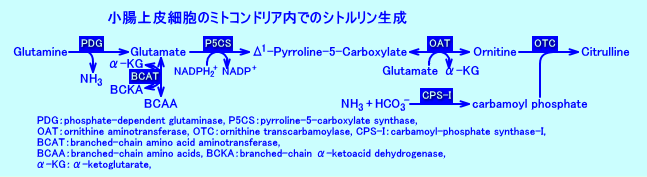
�@���T�F�����̏��זE�iabsorptive epithelial cells of the small intestine�j�̃~�g�R���h���A���ɂ́A�A�f�����ɕK�v�ȁAPDG�iphophate-dependent glutaminase�j�AP5CS�ipyrroline-5-carboxylate synthase�Fglutamate gamma-semialdehyde synthetase�j�AOAT�iornithine aminotransferase�j�ACPS-I�icarbamoyl-phosphate synthetase I�j�AOTC�iornithine transcarbamoylase�j�����݂���B
�@�����S���ɂ́A�H�a�R���̃O���^�~���_��A�H�a�R����z���t���̃O���^�~������PDG�ɂ�萶�������O���^�~���_��ABCAA����BCAT�ɂ�萶�������O���^�~���_���A���݂���B�����̃O���^�~���_��p���āA�����̏��זE�ł́AP5CS��OAT�iornithine aminotransferase�j�ɂ��A�I���j�`�����A���������B�I���j�`���́A�J���o�~�������_�i�������זE���ŁA�A�����j�A�Əd�Y�_�C�I���Ƃ���ACPS-I�ɂ�荇�������j�ƁAOTC�ɂ�茋�����āA�V�g�����������������B
�@�����Ő������ꂽ�V�g�������́A�z���t���ɕ��o����A�t���ŁA�A���M�j�������������B
�@�����ŁA�O���^�~���̒��f�́A37.9%���A�����j�A�ɁA27.6%���V�g�������ɁA24.4%���A���j���ɁA7.2%���v�������iproline�j�ɁA��ӂ����B�����ł́A�O���^�~���_��A�X�p���M���_�̒��f�́A�A�~�m��]�ڂɎg�p����A�قƂ�ǁA�A�����j�A�ɁA��ӂ���Ȃ��B
�@�����ł́A���f��ӂɂ��A�O���^�~���A�O���^�~���_�A�A�X�p���M���_�A�����āA���炭BCAA����AAST�iGOT�j�ɂ��A�A���j�������������B�̑��ł́A�A���j���ƃA�����j�A���A�喬����̓����������荞��ŁA�O���^�~������o�����i�A���j�����琶�������O���^�~���_�ƁA�A�����j�A��p���āA�O���^�~�������y�f�ɂ��A�O���^�~��������j�B�����́A���Ǔ��i�H���R���j�̃O���^�~����A���������̃O���^�~���𗘗p���A���ʂ̃A���j����A�����j�A���A�����ɕ��o����i�����ŃA���j�������A�̑��ŃO���^�~���ɂ��A�����ɖ߂��A���f�����T�C�N������j�B
�@�t���ɂ́A�A�f��H�̃A���M�j�m�R�n�N�_�����y�f�iargininosuccinate synthase�j�A�A���M�j�m�R�n�N�_�����y�f�iargininosuccinate lyase�j�����݂��Ă��āA�A���M�j�����A���������B�t���ɂ́A���ʂ̃A���M�i�[�[�����݂��Ă��āA�A�f�����������B
�@�Ȃ��A�A���M�j���́A�̑��ł́A�A���M�i�[�[�ɂ�蕪�������B�A���M�j���́A�A�f��H��H�ȊO�ɁANO��A�N���A�`���Ȃǂ̍����ɕK�v�B
�@���U�F���l�ł́A�P���ɁA4,150mg�̃A�����j�A���z�������ƌ����B
�@���V�F�A���j���́A�̑��ł́AALT�iGPT�j�ɂ��A�A�~�m���-�P�g�O���^���_�ɓ]�ڂ���A�s���r���_�ƂȂ�A�O���^�~���_�����������B�O���^�~���_�́A�~�g�R���h���A���ŁAm-AST�im-GOT�j�ɂ��A�A�~�m��]�ڂ���A�A�f��H�ɕK�v�ȃA�X�p���M���_�����������B�܂��A�O���^�~���_�́AGDH�i�̍זE�̃~�g�R���h���A���ɑ������݂���j�ɂ��A�_���I�E�A�~�m������A�A�����j�A�iNH4+�j���V�������B
�@���W�F�̑��ʼnߏ�ɐ������ꂽ�O���^�~���́A������t���ɗA������A�t�����O���^�~�i�[�[�ɂ��A�A�����j�A�ƃO���^�~���_�ƂɁA��������A���f�́A�A�����j�E���C�I���Ƃ��āA�A���ɔr�������B�O���^�~�i�[�[�ɂ�萶�������A�����j�A�́A�_����t�̈ێ��ɖ𗧂��Ă���B
�@�������A�A���ɔr������钂�f�̖�80���́A�A�����j�E���C�I���Ƃ��Ăł͂Ȃ��A�A�f�Ƃ��Ĕr�������B�A�f�̑啔���́A�̑��̔A�f��H�Ő��������B
�@�Q�l����
�@�E�n�[�p�[�E�����w�i����14�ŁA�O�Y�`���Ė�A�ۑP������ЁA�@1975�N�j�D
�@�E���H�[�g�����w�i�������w���l�A2003�N�A��S���j�D
�@�E��؍h��A���F�z�[�g�������w�@��R�Łi�������w���l�A2005�N�A��3���j�D
�@�EMalcolm Watford (2003) The Urea Cycle: Teachin Intermediary Metabolism In a Physiological Setting. Biochemistry and Molecular Biology Education 31, 289-297.
�@�E�͍����A�����T�F�����̗Տ�����2001/2002�i��]���A2001�N�j�D
�@�E�͍����F�ڂŌ��鏉���f�Â̌����v��ƌ��ʂ̓ǂݕ��i�G�X�A�[���G���A1997�N�j�D
�@�EMalcom Watford (2000) Glutamine and Glutamate Metabolism across the Liver Sinusoid. Journal of Nutrition. 130:983-987.
�@�EGuoyao Wu, et al. (2000) A Cortisol Surge Mediates the Enhanced Expression of Pig Intestinal Pyrroline-5-Carboxylate Synthase during Weaning. Journal of Nutrition 130:1914-1919.
�@�E�̎����f�Ã}�j���A���A���{��t��G���i���ʍ��j Vol.122, No.8 1999�N�D
�@�EGuoyao Wu (1998) Intestinal Mucosal Amino Acid Catabolism. The Journal of Nutrition 128:1249-1252.
�@�E���c��Y�A���F�I���j�`���g�����X�J���o�~���[�[�����ǂ̏o���O�f�f�ƈ�`�q���Á@�����ȁ@37:1475-1484, 1996.
�@�E�b�c���Y�F���H�����N�̌��_�@�i���o�ŁA1991�N���Ŕ��s�j�D
�@�E�c��M�v�F���炾�̓�������݂��ӂ̉h�{�w�@�^�J���o�C�I������Ёi2003�N�j�D
�@�E��䐴���A�a�c�`�Y�F�J���j�`����ӈُ�@�����ȁ@Vol.30 No.4�A363-368�A1989�N�D
�@�E�������u�F�i�[�V���OQ&A�@�S�ȂɕK�v�ȉh�{�Ǘ�Q&A�A������w�Ёi2005�N�j�D
�@�E�|��˘Y�F���^�����@�T���E�G�C�̐Z�������߁A���{�㎖�V��ANo.4309�i2006�N11��25���j�A93-94�ŁD
�@�EGerhard Gottschalk: Bacterial Metabolism�F���c�����A��ؗ�q�@��A�P�{���_�@�Ė�F�ۂ̑�ӁA�ߑ�o�Łi1980�N�j�D
�@�b�g�b�v�y�[�W�b�����ƌ����̊W�b�~�j��w�m���b�����w�̒m���b��w�̘b���b�����Ȏ����b�����̕s�v�c�b�����N�W�b