�@
�@ �@
�@ �@
�@�@�@�@
�@�d�q�`�B�n�Ǝ_���I�����_��
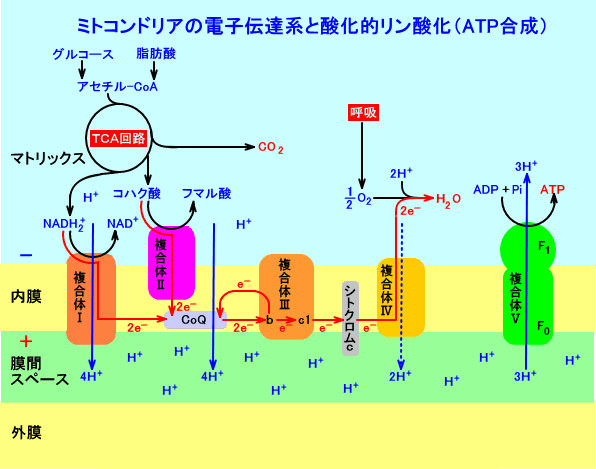
�@�~�g�R���h���A�imitochondria�j�ł́A�d�q�������ɓ`�B�����āA���f�C�I���iH+�F�v���g���j�����ԃX�y�[�X�i���ԍo�j�ɗA�����i�d�q�`�B�n�j�A������v���g���Z�x���z�i�d�ʍ��ApH���j��p���āA�����i���P�j�I��ATP�̍������s����i�_���I�����_���j�B�~�g�R���h���A���́A�d�ʂ�-180mV�ApH�́A8�B
�@�����S�ߒ��́A�ċz���irespiratory chain�j�ƌĂ�Ă���B
�@ �d�q�`�B�n�ł́A�d�q���A�_���Ҍ��d�ʂ̒Ⴂ�����iNADH2+�F�|0.315V�j����A���������iO2�F�{0.815V�j�ɁA�`�B�����B
�@�Ҍ����ꂽ�_�f�iO2�j�́A�v���g���iH+�j�ƌ������āA���iH2O�j�����������B
�@�~�g�R���h���A�̓����ɂ́A�T�̕����̂ƁACoQ�i��y�fQ�A���r�L�m���j�A�V�g�N����c�i�`�g�N���[��c�j�����݂���B
�@CoQ�́A�����������R�ɓ����܂��A�d�q��`�B������A�v���g���ԃX�y�[�X�ɋ��ݏo���B
�@
�@�M���ނł́AATP�̂W�O�`�X�O���́A�~�g�R���h���A�ł��_���I�����_�ɂ��A���������B
�@�~�g�R���h���A�́AATP���������čזE�̐����ێ����邾���łȂ��A�A�|�g�[�V�X�ɒ��S�I�Ȗ������ʂ����āA�זE�̎��������䂵�Ă���B
�@�d�q�`�B�n�ł́ANADH2+��FADH2+�̐��f���_�������B
�@NADH2+�́A�Ő�������~�g�R���h���A���ɊҌ����ʂ��A�����ꂽ��ATCA��H�ŃA�Z�`��-CoA�i��A���b�_����-�_����A�A�~�m�_��ӂŐ��������j����ӂ��ꐶ�����ꂽ��A���b�_�̃�-�_���ɂ�萶�������B
�@FADH2�́A���b�_�̃�-�_���ɂ��A���������B
�@�d�q�`�B�n�ł́A�_���Ҍ������ɂ��A�d�q�́A�_���Ҍ��d�ʂ̒Ⴂ�����iNADH2+�F�|0.315V�j����A���������iO2�F�{0.815V�j�ɁA�`�B�����B
�@�d�q�`�B�n�ł́ANADH2+�Ƃ��đ�����ꂽ�A�����⎉�b�_�R���̊Ҍ����ʁireducing equivalents�j���A�_�f�ɓ`�B���A�������������iNADH2+��FADH2+���f���_�������j�B
�@�d�q�`�B�n�ł́ATCA��H�i�N�G���_��H�j�����ŎY�����ꂽNADH2+�i���Q�j��A�R�n�N�_���A�_������A�d�q�ie-�j�����o�����B
�@���o�����d�q�ie-�j���A�����ɑ��݂��镡���̊Ԃ��A�`�B�����B
�@�d�q�`�B�ɔ����A���f�C�I���iH+�F�v���g���j���A���d�ʂɋt����āA���ԃX�y�[�X�ɗA�������B���̌��ʁA�~�g�R���h���A���i�}�g���b�N�X�j�́A�d�ʂ�-180mV�ApH�́A8�ƂȂ�B
�@�`�B���ꂽ�d�q���A�_�f���q���Ҍ������A����ɐ��f�C�I���ƌ��������āA�������������B
�@���D������I�iNADH dehydrogenase�ANADH-CoQ reductase�j
�@������I�́AFMN�AFe-S�����B
�@�~�g�R���h���A���}�g���b�N�X�ł́A�s���r���_��TCA��H�ł̕�����A���b�_����-�_���ŕ��o�����A���f���q���A�_���Ҍ������̕�y�f�ł���ANAD+�inicotinamide adenine dinucleotide�j���Ҍ����ANADH2+�����������B
�@�Ȃ��A���ŎY�������NADH2+���A�זE���]������A�~�g�R���h���A�̃}�g���b�N�X���i�}�g���b�N�X�j�ɓ���ɂ́A�����S�_-�A�X�p���M���_�V���g���i�̑��A�S���A�t���Ȃǁj��A�O���Z�������_�V���g���i�ؓ��ȂǁA���R�j���A�g�p����B
�@������I�́ANADH2+�i���S�j�̂Q�P�̐��f�C�I���iH+�j�ƂQ�P�̓d�q�ie-�j���ACoQ�i��y�fQ�A���r�L�m���j�ɓ`�B����B
�@������I�ƕ�����II�́A�E���f�y�f�����́B
�@ �Ȃ��A������I�ɂ́A�v���g���|���v�̍�p������A�v���g���iH+�F���f�C�I���j���A�}�g���b�N�X����A�~�g�R���h���A�������o�āA���ԃX�y�[�X�ɋ��ݏo���ƌ�����������B���̏ꍇ�ANADH2+�̓d�q�P�i�Q�P�̓d�q�j����A�S�P�̃v���g���iH+�j���A���ԃX�y�[�X�ɋ��ݏo���A�ƍl�����Ă���i���T�j�B
�@���D������II�isuccinate dehydrogenase�j
�@������II�́ATCA��H�i�N�G���_��H�j�́A�R�n�N�_�E���f�y�f�Ɠ����ŁAFAD�ƁAFe-S�����B
�@������II�́A�R�n�N�_����A�Q�P�̐��f�C�I���ƂQ�P�̓d�q���AFAD����āACoQ�i���r�L�m���j�ɓ`�B���A�Ҍ��^��C��QH2�i���r�L�m�[���j���o����B
�@�R�n�N�_�{FAD���t�}���_�{FADH2
�@FADH2�{C��Q��FAD�{C��QH2�i�d�q�́AFAD��Fe3+S��C��Q�Ɠ`�B�����j
�@���b�_����-�_�������ۂɁAFADH2���Y������i��-�_���ł́ANADH2+�����������j�A�d�q�`�B�t���r���^���p�N�� �iETF�Felectoron transfer flavoprotein�j���o�āA�d�q���`�B�����B
�@�V�g�N����bc1�����́A�V�g�N�����Ҍ��y�f�Ƃ��Ă��B
�@�Ҍ��^CoQ�iC��QH2�A���r�L�m�[���j�́A������III�̃V�g�N����b�ɁA�Q�P�̓d�q��`�B���A�Q�P�̐��f�C�I���ԃX�y�[�X�ɋ��ݏo���A�_���^C��Q�ɖ߂�B
�@���̍ہA�Q�P�̓d�q�̓��A�P�P�́A������III�̃V�g�N����c1���o�āA�~�g�R���h���A�����̖��ԃX�y�[�X���ɑ��݂���V�g�N����c�ɓ`�B�����B�����P�P�̓d�q�́A�V�g�N����b�����ړ����āA�_���^C��Q�ɓ`�B����A��d�q�Ҍ�����B���̊Ҍ��^C��Q�iC��Q�E-�̃��W�J���j�́A�}�g���b�N�X������Q�P�̐��f�C�I������荞�݁A���ԃX�y�[�X�ɐ��f�C�I�������ݏo���Ƃ����iQ�T�C�N���j�B
�@���D�V�g�N����c
�@�V�g�N����c�i�`�g�N���[��c�j�́A�~�g�R���h���A�����̖��ԃX�y�[�X���ɕ\�݂���n���`�����B
�@�V�g�N����c�́A�w���S��F�f�����Ɏ����Ă���B
�@������III����P�P�̓d�q�����A������IV�ɓ`�B����B
�@Fe2+�i�Ҍ��^�j��Fe3+�i�_���^�j�{e-
�@�V�g�N����c�i�`�g�N���[��c�j�́A�A�|�g�[�V�X�̍ۂɁA�~�g�R���h���A������o�����B
�@���D������IV�icytochrome oxidase�j
�@�V�g�N����c�I�L�V�_�[�[�A�V�g�N����aa3�����̂Ƃ��Ă��B
�@�V�g�N����c����A������IV�ɓn���ꂽ�d�q�ɂ��A�_�f���q�iO2�j���l�d�q�Ҍ�����ifour-electron reduction�FFea33+�|O-�|O-�|CuB2+�j�A����ɁA�}�g���b�N�X�����荞�܂ꂽ���f�C�I���iH+�j�ƌ������āA���iH2O�j�����������B
�@���̍ہA�ꕔ�̎_�f���q�iO2�j�́A��d�q�Ҍ��ione-electron reduction�j����āA�X�[�p�[�I�L�V�h�iO2-�j�ƌ��������_�f�ireactive oxygen species�FROS�j�ɂȂ�i���U�j�FO2�{e-��O2-�B
�@�V�A���C�I���iCN-�j�A�������f�iH2S�j�́A������IV��j�Q���A�Ő��������B
�@���̂悤�ɁANADH2+����́A�d�q���ANAD+��������I�iFMN��Fe3+S�j��C��Q��������III�i�V�g�N����b��Fe3+S���V�g�N����c1�j���V�g�N����c��������IV��O2�ƁA�_���Ҍ��d�ʂ̒Ⴂ�������獂�������̕��ɁA�`�B�����i���V�j�B
�@�܂��A���̂悤�ɁA�d�q�`�B���ꂽ�d�q�ɂ��A�_�f�iO2�j���Ҍ�����āA���iH2O�j�ɂȂ�B�ċz�Ŕr�o�������_���Y�f�iCO2�j�̎_�f���q�iO�j�́A���i�O���R�[�X�FC6H12O6�Ȃǁj�R���ł���A��C���̎_�f�iO�j�R���ł͂Ȃ��i���W�j�B
�@C6H12O6�{6O2��6CO2�{6H2O
�@�O���R�R���`�R�C�h�i���t�玿�z�������j�̃n�C�h���R�[�`�]���ihydrocortisone�j�A�v���h�j�]�����iprednisolone�j�A
�f�N�T���T�]���idexamethasone�j�́A������IV��j�Q�iinhibit�j����B
�@IFN-���́A�������}�N���t�@�[�W�ɁAiNOS�iinducible nitric oxide synthase�j��U�����A�Y�������NO�́A������IV��}������B�Ȃ��AIFN-���́A������I�̃T�u���j�b�g�ł���NDUFV1�̔�����}������B
�@�����Ɋ֗^����PGE2�̌����ɂȂ��A���L�h���_�́A�����̂h�ƕ����̂h�h�h���A�I��I�ɑj�Q���A�~�g�R���h���A���ߎ_�����f�ihydrogen peroxide�j�Y�����A�L�ӂɍ��߂�B�s�O�a���b�_�̕����A�O�a���b�_���A�j�Q��p�������B���̂悤�ɁA�����ƁA�_���I�����_���iOXPHOS�j�Ƃ̊Ԃɂ́A�֘A������iVane���j�B
�@�Q�D�_���I�����_���imitochondrial oxidative phosphorylation�FOXPHOS�j
�@��L�̂悤�ɁA�d�q�`�B�n�̉ߒ��ŁA�}�g���b�N�X�����疌�ԃX�y�[�X���ɐ��f�C�I�����A�������i�O���́A���f�C�I���߂�����̂ŁA�}�g���b�N�X�̐��f�C�I���Z�x���Ⴍ�Ȃ�j�B
�@���̌��ʁA�v���g���iH+�j�Z�x���z���A�~�g�R���h���A�������u�ĂāA���ԃX�y�[�X�ƃ}�g���b�N�X�̊ԂɁA������B�Ȃ��A�v���g���iH+�j�́A�~�g�R���h���A�O�������R�ɒʉߏo����̂ŁA���ԃX�y�[�X�́A�זE���]���ƁA�g�|���W�[�I�ɁA�����ł���B
�@���D������V�iF1F0-ATPase�AATP�����y�f�AH+-ATPase�j
�@�~�g�R���h���A�̖��ԃX�y�[�X������AATP�����y�f�ł��镡����V���o�āA�}�g���b�N�X���ɂR�P��H+�������ۂɁA�P�P��ATP�����������B������V�́A���[�^�[���q�ŁA�P�b�ԂɂP�O�O��ȏ�A��]�i�����j���āAATP���������Ă���B
�@ATP��ADP�́A�~�g�R���h���A������ʉߏo���Ȃ��̂ŁA�����E���o�́AATP/ADP�g�����X���J�[�[�ɂ���čs�Ȃ���Ƃ����B
�@�v���g���Z�x���z�𗘗p���āAATP�����y�f��ATP����������d�g�݂́A�A�����������ɍۂ��āA�t�Α̂̃`���R�C�h�ŁAATP�����������������_���iphotophosphorylation�j�̎d�g�݂ɋߎ����Ă���i���W�j�B
�@�R�D�ċz���̊���
�@�~�g�R���h���A�̌ċz���i�d�q�`�B�n�Ǝ_���I�����_���j�̊����i�����j�́A�P���i���v�^���Ȃǁj�ɂ���āA�K���I�ɑ��������B
�@���i�A�]��^�������Ă��Ȃ��l�́A�~�g�R���h���A�̌ċz���̊����i�_�������j���ア�B���i�A�]��^�������Ă��Ȃ��l�́A��-�_�������i���b�_����-�_���ŕ�������\�́j���A���Ɏア�B��-�_�������́A�����b�H�i�������b�_�j�����Ȏ����������i�̐ێ�j�ɂ���āA���������i�y�f�̓]�ʂ��A�U�������j�B��-�_�������́A�^���Ȃǂ̌P���ŁA��������ɂ́A�������V�����b�_�Z�x�������Ȃ���i�ጌ�����j�ɍs���̂��ǂ��ƌ����B
�@�S�DUCP
�@UCP�iuncoupling protein�j�́A�~�g�R���h���A�ɂ���E����`�����ŁA�l�X�Ȏh���ɂ��M�����B
�@�H���R���i�����⎉���R���j�̃G�l���M�[��iNADH2+�Ȃǁj�́A�~�g�R���h���A�ŁAATP�i���G�l���M�[�����_�������j�ɕϊ������B�G�l���M�[��̃G�l���M�[�̓��AATP�ɕϊ����������i�ϊ������j�́A��������40�����x�ƌ����A�����ȏ�́A�M�ɕϊ������i�M�Y���j�B�Y�������M�́A�̉����߂Ɋ�^����B
�@��ӂ̊����ȑ���i�̑��Ȃǁj�ł́A�����Ԃ̃G���W�����M�����悤�ɁA�M�������Y�������B�����̋@�\����肭�쓮���Ȃ��ƁA�M�����ȂǁA�T�M�ɂ���Q��������B
�@�P�j�DUCP1
�@UCP1�́A���F���b�g�D�ɓ��ٓI�ɑ��݂��A�M�Y���@�\���ʂ����B���F���b�g�D�́A��ӂ邦�M�Y���ɂ��A�M�Y�����s���i���i�́A�ӂ邦�M�Y���ɂ��A�M�Y�����s���j�B
�@UCP1�́A�M�Y���\�͂������BUCP1�ɂ��M�Y���́A�m���A�h���i�����ɂ�董�i��������F�m���A�h���i�����ɂ��h���ɂ���ӗʂ���10�{�ɑ������A�זE�������3�i�m���b�g�ɑ�������M���Y������F�g�D1kg�������300���b�g�i300���b�g/kg�j�̔M���Y������i�M����������b����́A4.1���b�g/1kg�j�BUCP1�����������}�E�X�́A����ϐ����ቺ����FUCP1�����������}�E�X�́A�₦���ɂȂ�A�ʏ�̎������ł��A�������ǂ����k�����A�M���U���A�}������BUCP1�����������}�E�X�́A�����b�H���ƁA����Ƌ��ɁA�얞�ɂȂ�i�C���X������R�����j�B
�@
�@�Q�j�DUCP3
�@UCP3�́A���i�ɔ������Ă���iUCP3���A�M�Y���\�͂�L���邩�A�c�_��������Ă���j�B
�@UCP3�̔����ʂ́A�^���ɂ�葝�����A�V���Ƌ��Ɍ�������B
�@UCP3��`�q��UCP1��`�q�̔����ʂ́A�b��B�z�������̃g�����[�h�T�C���j���iT3�j�ɂ��A������������iT3�́A���i�����F���b�g�D�̔M�Y���̒��߂Ɋ֗^���Ă���\��������j�B
�@�T�D�b��B�z������
�@�b��B�z�������ithyroxin�j�́A�_���I�����_�����A���i������B
�@�b��B�@�\���i�ǂł́A�g�����[�h�T�C���j���itriiodothyronine�FT3�j�ɂ��A�S���ł�ATP����������邪�AATP�����i�_���I�����_���ɂ�萶�������j�ƁA�ؓ����k�iATP��������j�����A�M�Y���ithermogenesis�j�̕��ɁA���d�ʁi���̃G�l���M�[�j���A�g�p�����B
�@�~�g�R���h���A�ɂ́A�b��B�z�������ƍ����e�a���Ō��������e�̂����݂���B
�@�b��B�z�������́A���b�g�̐t���ŁA�זE���̒`������āA�~�g�R���h���A�̒`�������𑣐i������B
�@�b��B�z�������́A���b�g�̐S�؍זE�ŁA�זE���ɂ����āA�}���������̗ʂ������A�~�g�R���h���A�O���ɑ��݂���MAO�i���m�A�~���I�L�V�_�[�[�j������}������B
�@�U�D�V�g�N����c �I�L�V�_�[�[������
�@�V�g�N����c �I�L�V�_�[�[�icytochrome c oxidase�FCCO�j���A��V�I�Ɍ��������V�g�N����c �I�L�V�_�[�[�����ǁicytochrome c oxidase deficiency�j�ł́A�������������璘�����؋ْ��ቺ�A�`���A�A�A���Ȃǂ�������B
�@�~�g�R���h���A�Ɉُ킪����ؐ��ۂ́A���w�����������imodified Gomori trichrome���F�j�ł́A�������Ԑ����A�����ڂ�ڂ�Ƃ�������������iragged-red fiber�j�B
�@�d�q�����������ł́A�~�g�R���h���A�̌`�Ԉُ킪������F�~�g�R���h���A�́A����ŁA�N���X�e�����G�ɑ������Ă���B
�@�~�g�R���h���A�̌`�Ԉُ�́ACPT�����ǁA�J���j�`�������ǁA�d�q�`�B�n�ُ̈�iCCO�����Ǔ��j�ŁA������B
�@�V�D���̑�
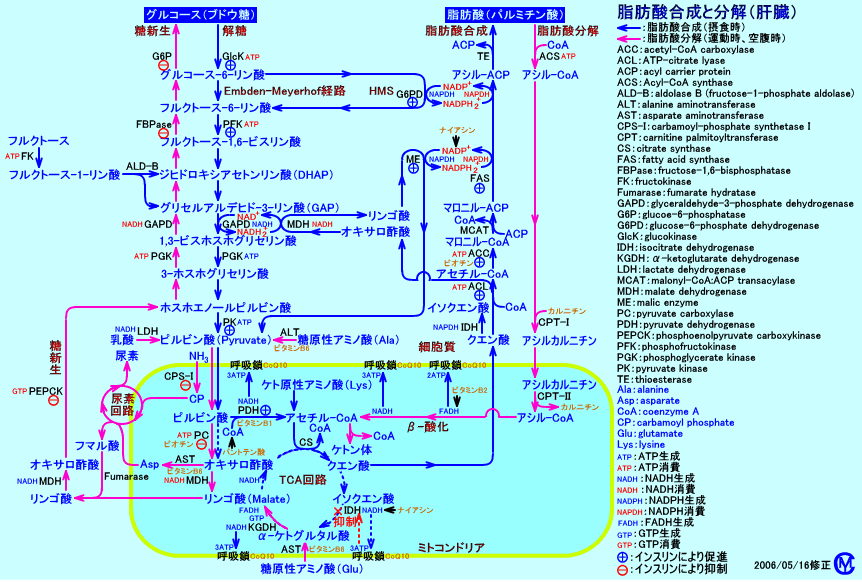
�@�E�ہi�咰�ہj���A�O���R�[�X�Ɩ��@���ނ�p���āA�זE����1g����������ɂ́AATP�͋͂�34.8mmol�K�v�Ƃ���ɉ߂��Ȃ��iATP��1mol�p���āA�זE��30�������������j�B
�@�זE�����̍����ɗ��p�����34.8mmol��ATP�̓��A19.1mmol�i56���j�̓A�~�m�_���d�����Ē`��������������̂ɁA���p�����B
�@�O���R�[�X����A�~�m�_������̂ɗv����ATP�ʂ́A1.4mmol�i��1/10�ʁj�ƌ�����B
�@���P�F�d�q�`�B�n�Ǝ_���I�����_���́A�u�����v�icoupling�j���Ă���̂ŁA�_���I�����_����ATP����������Ȃ��ƁA�d�q�`�B���N����Ȃ��B
�@2,4-�W�j�g���t�F�m�[���́A�E�����܁F2,4-�W�j�g���t�F�m�[���́A�~�g�R���h���A�̓�����ʉߏo����B2,4-�W�j�g���t�F�m�[���́A���ԃX�y�[�X�ŁA���f�C�I���i�v���g���FH+�j�ƌ������āA��������𗣌^�́A�}�g���b�N�X�Ɉڍs����B���̌��ʁA�d�q�`�B�Ő�����A�v���g�����z���Ȃ��Ȃ�AATP�������j�Q�����B
�@�܂��A�V������~�������ł́A���F���b�g�D�̍זE�̃~�g�R���h���A�ŁA�v���g�����A������V�iF1F0�j�Ƃ͈قȂ�v���g���`���l����ʉ߂��āA�}�g���b�N�X�ɗ����i�E��������j�̂ŁAATP�������A�M�Y���i�M�����j���N����B����ɂ��A�����_�o���h�������ƁA���b�������N����A�V���������b�_���A���F���b�זE�̃~�g�R���h���A�����ɑ��ʂɑ��݂���Athermogenin�ƌĂ��E����`�����iuncoupling protein�FUCP�j���J���A���d�ʁi�~�g�R���h���A�̃v���g���̔Z�x���z�j����������BUCP�́A�T�[���j�Q���i�M�����^���p�N�j�Ƃ��Ă��B��������ƁAATP�������s���Ȃ����A���b�_�ȂǁA��̎_�������������i���āA�M�Y�����N����i���b�_����-�_�������i����A�M�G�l���M�[�����o�����j�B���F���b�זE���j�ꂽ�����́A�C�����ቺ�����ۂ̑̉��ێ��@�\���A�ቺ����B�܂��A���F���b�זE���j�ꂽ�����́A�i�M�Y���ɂ��J�����[����ቺ���āA�j�̎��b�ʂ�̏d����������B
�@�̓��̑�ӂŐ����鐶�̃G�l���M�[�̓��A�~�g�R���h���A��ATP�ɕϊ������i�ؓ��Ȃǂ̎d���ɗ��p�����j���́A15�`20���ƌ�����i�ő�ł�45�����x�j�B�c��̐��̃G�l���M�[�́A�M�G�l���M�[�ɕϊ������i�M�Y���ɗ��p�����j�B
�@�ʏ�́A�̓��̑�ӂŐ����鐶�̃G�l���M�[�́AATP�����ɗ��p����A�Ȃ�ׂ��A�M�Y���ɂ́A���p����Ȃ����A���⎞�Ȃǂ́A�̉���ቺ�����Ȃ��悤�ɁA�M�Y����������ׁAATP�������]���ɂ��āA���̃G�l���M�[��M�G�l���M�[�ɕϊ�����BATP�́A�~�g�R���h���A�̌ċz���Ő�������邪�A�E�������邱�ƂŁAATP������}�����A�M�Y����������B
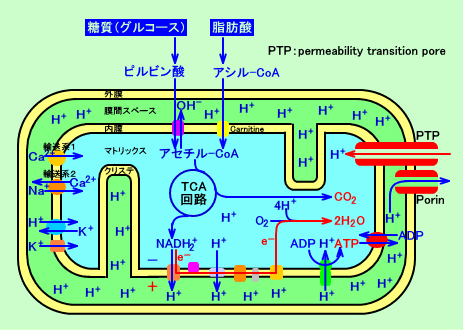
�@�T���`���_�Ȃǂ�NSAIDs�́APTP�ƌ����A�~�g�R���h���A�̓����ƊO�����ђʂ��錊�\�����A�J�������āA�v���g�����A�~�g�R���h���A���ɗ��������A���d�ʂ�ቺ�����A�A�|�g�[�V�X�ŁA�זE�����ł����Ă��܂��B
�@�A�|�g�[�V�X����N����PTP�̊J���iinduction�j�́ACa2+�Ɉˑ������B���̗��R�́ABernardi�̎������ʂ���APTP�̊J���́A���d�ʁithe proton electrochemical gradient�F����H+�j�ɂ�萧�䂳��Ă��āACa2+�iCam�̑����j���A�~�g�R���h���A���ŁA���d�ʁi����H+�j��ω������āAPTP�̊J���iinduction�j�������N�������߂ƁA�l������B
�@Trost���̎������ʂł́A�זE�OCa2+�Z�x�iCao�j�������ƁA�T���`���_�̓Ő��i�~�g�R���h���A��Q��p�j���A���������B�זE�O�iextracellular�j�̃J���V�E���C�I���Z�x�iCa2+�Z�x�j�������ƁA�T���`���_�́A�Ő��i�~�g�R���h���A��Q��p�j���A���������B�J���V�E���h�R��p�̂����܁iverapamil�Adiltiazem�Achlorpromazine�Anifedipine�Anisoldipine�j�́A�T���`���_�̓Ő��i�~�g�R���h���A��Q��p�j���A�j�Q�Ȃ����y���������B
�@���R�FNADH2+�́ANADH�{H+�̂��ƁB
�@���S�F�O���Z�������_�V���g���́A�~�g�R���h���A�����̖��ԃX�y�[�X���i���ԕ����j�ɑ��݂��A�ȉ��ɏq�ׂ�悤�ȂR�X�e�b�v�̔����i�d�g�݁j�ŁANADH2+���A�~�g�R���h���A���ɁA�A������B
�@�P�j�D�זE���]���i�T�C�g�]���j��NADH2+�̓d�q�iH�j�́A�O���Z���[��-3-�����_�f�q�h���Q�i�[�[�i3-�z�X�z�O���Z���[���f�q�h���Q�i�[�[�j�ɂ��A�W�q�h���L�V�A�Z�g�������_�iDHAP�j���Ҍ����ANAD+�ƁA�O���Z���[��3-�����_�i�O���Z���[��-3-�����_�A��-�O���Z�������_�j���A���������BNAD+�́A�n�ŁA�ė��p�����B
�@�Q�j�D�O���Z���[��3-�����_�̓d�q�iH�j�́A�~�g�R���h���A�����̖��ԃX�y�[�X���i���ԕ����j�ŁA�t���{�v���e�C���f�q�h���Q�i�[�[�ɂ��AFAD+���Ҍ����AFADH2+���A��������A�W�q�h���L�V�A�Z�g�������_�ɖ߂�B
�@�R�j�DFADH2+�́A�~�g�R���h���A�����ŁA�d�q�iH�j���A�d�q�`�B�n�ɑ��荞�݁AFAD+�ɖ߂�B
�@���T�F�v���g���|���v�́A�~�g�R���h���A�����ɑ��݂��Ă���B�v���g���́A�_���^�v���g���|���v�ł́A�v���g���|���v�̃}�g���b�N�X���̃A�~�m�_�����Ɍ������Ă���B
�@�_���^�v���g���|���v�́A�d�q�`�B�ŊҌ������ƁA�\���i�R���t�H���[�V�����j���ω����āA�v���g�������������𗣊�i�A�~�m�_�����j���A�זE���]�����Ɍ�����ς��āA�v���g�����A���ԃX�y�[�X�ɉ𗣂���B�Ҍ��^�v���g���|���v�́A�Ď_�������ƁA�\�����ω����āA�}�g���b�N�X���Ɍ�����ς���B
�@���̂悤�ɂ��āA�d�q�`�B�ŁA�_���^�v���g���|���v���A�Ҍ��^�ɍ\����ω����邱�ƂŁA�}�g���b�N������A���ԃX�y�[�X�ɁANADH�̓d�q�P�Γ���A�S�P�̃v���g�����A���ݏo���i�A������j�ƌ����B�@
�@���U�F������IV�ɓn���ꂽ�d�q�ɂ��A�_�f���q�iO2�j���Ҍ������ۂɁA�ꕔ�̎_�f���q�́A��d�q�Ҍ�����A�X�[�p�[�I�L�V�h�iO2-�j�A�ߎ_�����f�iH2O2�j�A�q�h���L�V�����W�J���iHO�E�j�Ȃ������_�f�iROS�j���A�������Ă��܂��B
�@�����_�f�́A�����̂h�h�h�ł��A�d�q�i�v���g���j���ACoQ�iubiquinone�j�A�V�g�N����b�A�V�g�N����c1�̊Ԃ��A�s��������Ԃɔ�������B������IV�Ŕ������銈���_�f�̗ʂ����A�����̂h�h�h�Ŕ������銈���_�f�̗ʂ̕����A�����ƌ����B
�@�����̂h�ł��A�����_�f����������B
�@���V�F�_���Ҍ��d�������̕����́A�d�q�e�a�͂����f���Ⴍ�A�d�q��^���Ղ��B
�@�_���Ҍ��d�ʂ����̕����́A�d�q�e�a�͂����f��荂���A�d�q�����Ղ��B
�@�d�q�`�B�n�ł́ANADH2+����1/2O2�ɁA�Q�P�̓d�q���`�B����邪�ANADH2+�i-0.315V�j��O2�i+0.815V�j�Ƃ̊Ԃɂ́A1.13V�i�{���g�j�̓d�ʍ�������B�d�q1mol���A1V�̓d�ʍ��̊Ԃ��ړ�����ƁA-96.5kJ�̓d�C�G�l���M�[�i���R�G�l���M�[�F��G�j���A���o�����̂ŁA���R�G�l���M�[�ω���Go'�́A-222kJ/mol�B
�@�d�q�́A�d�q�`�B�n�ł́A �G�l���M�[���ʂ������Ƃ���i�_���Ҍ��d�ʂ��Ⴂ�����FNADH2+�j����A�Ⴂ�Ƃ���i�_���Ҍ��d�ʂ����������FO2�j�ւƗ���A���̍ۂɕ��o�����G�l���M�[�i��Go'�j���A�v���g���̗A���ɗp������B
�@�W���_���Ҍ��d�� �@�_���Ҍ��n �@Eo' (V) �@�s���r���_���|�_�{CO2 �@-0.70 �@�|�_�{3H+�{2e-���A�Z�g�A���f�q�h�{H2O �@-0.581 �@H+�{e-��1/2H2 �@-0.421 �@NAD+�{H+�{2e-��NADH �@-0.315 �@FAD�{2H+�{2e-��FADH2�i�V���̕�y�f�j �@-0.219 �@�s���r���_ + 2H+ + 2e-�� ���_ �@-0.185 �@�I�L�T���|�_�{2H+�{2e-�������S�_ �@-0.166 �@FAD�{2H+�{2e-��FADH2�i�t���{�^���p�N���j �@-0. �@���r�L�m���{2H+�{2e-�����r�L�m�[�� �@+0.045 �@1/2O2�{2H+�{2e-��H2O �@+0.815 �@�����̃C�I�����X���́A�������f���L���Ă���A�_���Ҍ��d�ʂ̑召�̏����A�����Ă���B
�@�C�I�����X���F�݂����iK�j���iCa�j�ȁiNa�j�A�܁iMg�j���iAl�j���i:Zn�F�����j�āiFe�F�S�j�ɁiNi�j����iSn�j�ȁiPb�F���j�A�ЁiH�j�ǁiCu�F���j���iHg�F����j����iAg�F��j�iPt�F�����j���iAu�F���j
�@���W�F�M�u�Y�iGibbs�j�̎��R�G�l���M�[�́ACO2�F-394.36kJ mol-1�AH2O�F-237.13kJ mol-1�AC6H12O6�F-910kJ mol-1�Ȃ̂ŁA
�@C6H12O6�{6O2��6CO2�{6H2O
�@�̔����ł́A2880kJ ��������B
�@���̂́A�����ǂ�ATP�����邪�A������ӂŐ����鑍�G�l���M�[�̂����AATP�ɕϊ��o����̂́A45���ȉ��ƌ�����F�M�@���i���C�@�֎ԁj�̌������A10�����x�ł��邱�Ƃ��l����ƁA���̂�ATP�ւ̃G�l���M�[�ϊ������́A���ɗǂ��i�ŋ߂ł́A�M�����́A�K�\�����G���W����25�����x�A�f�B�[�[���G���W����30�����x�ƌ�����j�B�c���55���ȏ�̃G�l���M�[�́A�M�G�l���M�[�ɕς��A�̔M�Ƃ��āA�̕\�ʂ���g�U����Ă��܂��B�H�����̃G�l���M�[�ʂ�100���Ƃ���ƁA95�������R�G�l���M�[�i�G�l���M�[�ʂ�5���́A�G���g���s�[�ω��ɂ��M�ʂɕς��j�Ƃ��Ď��o����A�ő�45�����AATP�ɕϊ������BATP�ɕϊ����ꂽ���R�G�l���M�[�́A�������A�i�ؓ��j�^���A�\���A���Ȃǂ̐��̂̎d���ɁA�g�p�����B�T�O����ATP�ɕϊ�����Ȃ��������R�G�l���M�[��A���̂̎d���Ɏg�p���ꂽATP����A�M�ʂ��A������i�M�Y�����N����j�B
�@�Ȃ��AATP�̑�ӂł́A���L�̂悤�Ȏ��R�G�l���M�[����������B
�@MgATP2- + H2O �� MgADP- + Pi2- + H+ �F-30.3 kJ mol-1
�@ATP4- + H2O �� AMP2- + PPi3- + H+ �F-37.4 kJ mol-1
�@ADP3- + H2O �� AMP2- + Pi2- + H+ �F-36.3 kJ mol-1
�@AMP2- + H2O �� adenosine + Pi2- �F- 9.6 kJ mol-1
�@PPi3- + H2O �� 2 Pi2- + H+ �F-33.4 kJ mol-1
�@
�@1���Q�C�T�T�OKcal�������q�g�ł́AATP�i���q�ʁ���T�O�O�A���������G�l���M�[���V�D�RKcal/mol�j�����̕ϊ��������T�O���Ƃ���ƁA1���ɂP�V�Tmol�i�W�Vkg�j��ATP����������邱�ƂɂȂ�B�̓��ɑ��݂���ATP�ʂ́A��P�O�Og�Ȃ̂ŁAATP�́A�����E�������J��Ԃ��Ă���B
�@�q�g�̑̓��̊̑��⍜���ł́A�������ꂽATP�̂X�O���ȏオ�A�̂��\������`�����A�j�_�A�����ނȂǂ̉��w�����ׂ̈ɁA�g�p�����B
�@���W�F�A���ɂ́A�������m�������e�̂����݂���B
�@����e�̂ɂ́A�ԐF���i�Ԃ����FR�j�����m����t�B�g�N�����A�F���i�����j�����m����t�H�g�g���s���A�N���v�g�N���������݂���B
�@�P�j.�t�B�g�N����
�@�t�B�g�N�����́A�ԐF���i�Ԃ����FR�j�����m���āA����𑣐i����B�ԐF���Ă���A���ԐF���iFR�Ffar red�j�Ă�ƁA�����}������i�t�B�g�N�����́A�ԐF�������m���āA����𑣐i�����A���ԐF�������m���āA�����}������j�B
�@�t�B�g�N�������A�ԐF�������m����ƁA�s�̐L�т��������A�t���L��������B�t�B�g�N�������A���ԐF�������m����ƁA�s��L��������B
�@�t�B�g�N����A�́A�ア����g���ɊW�Ȃ����m���Ă���B�t�B�g�N����B�́A���̔g�����������Ă���B�t�B�g�N����B���A�ԐF�������m���A�����A�s�̐L���Ɋ֗^���Ă���B�t�B�g�N����B�́A�ԐF�������m����ƁA�j���Ɉڍs���A�M����`�B����B����e�́i����e�`�����j�̃t�B�g�N����B�́A�ԐF���ɂ�芈���^�ɕω����i����𑣐i����j�A���ԐF���ɂ��s�����^�ɕω�����i�����}������j�B
�@�ԐF���iR�j�����ԐF���iFR�j���ƁA�A���̐L�����}������A����i�ԉ蕪���j�����i�����i���K�N�[���j�B
�@�ԐF���iR�j�����ԐF���iFR�j���ƁA�A���̐L�������i����A���肪�}�������i�|�I�p�I�j�B
�@�Q�j.�t�H�g�g���s��
�@�t�H�g�g���s���́A�F���i�����j�����m����B�t�Α̂́A�זE���ŁA�����ɓ����Ă������A�����������m����ƁA�זE���ŁA���Ӂi�זE���߂��j�ɓ����čs���A�����キ�Ȃ�ƁA�זE�̒��S�ɖ߂��ė���i����ʉ^���j�B
�@�t�H�g�g���s��2���A�F�������m����ƁA����ʉ^�����N����B
�@�R�j�t�B�g�N����3
�@�V�_�́A�t�B�g�N����3��L���Ă��āA�t�B�g�N�����̂悤�ɐԐF���i�Ԃ����FR�j�ƁA�t�H�g�g���s���̂悤�ɐF���i�����FB�j�̗��������m����B
�@�ԐF���iR�j���F���iB�j���ƁA�t�ؗނ̐L�������i����i�̑��z�F�āj�B
�@�ԐF���iR�j���F���iB�j���ƁA�����ԙ��ނ̊J�Ԃ����i����i�Ԃ̑��z�F�~�j�B
�@���z���́A�Ăɂ́A�g���̒Z���F�̌��̊������������i�̑��z�j�A���ɁA�~�ɂ́A�g���̒����ԐF�̌��̊�������������i�Ԃ̑��z�j�B�]���āA�A���́A�ď�ɂ́A�̑��z�ɂ��L�����i�s���L�т�j�A�~�ꂩ��́A�Ԃ̑��z�ɂ��ԉ��t����B
�@�ԐF�����_�C�I�[�h�̐ԐF���iR�j��t�Αf�i�N�����t�B���j�ɏƎ˂���ƁA�����������i����A�H���|�̌s��t���L������i�ԐF���́A�A����L��������j�B
�@�F�����_�C�I�[�h�̐F���iB�j��t�Αf�i�N�����t�B���j�ɏƎ˂���ƁA�A���̌s�������Ȃ�����A�t�������Ȃ����肷��i�F���́A�A���̗t��s������j�B
�@�����́A300nm���x�̔g���̌��i�����O���j���D�ނ̂ŁA�U�����́A���̔g���̌��Œ���������B
�@�Q�l����
�@�E�n�[�p�[�E�����w�i����14�ŁA�O�Y�`���Ė�A�ۑP������ЁA�@1975�N�j�D
�@�E���H�[�g��b�����w�i�������w���l�A��P�ő�S���A2003�N�j.
�@�E��؍h��A���F�z�[�g�������w�@��R�Łi�������w���l�A2005�N�A��3���j�D
�@�E�V���v�������w�i��]���A������4�ŁA2003�N�j�D
�@�E��ؑO�A���F�Տ������w�@�㊪�i��R���A1975�N�j�D
�@�E�c��M�v�F���炾�̓�������݂��ӂ̉h�{�w�@�^�J���o�C�I������Ёi2003�N�j�D
�@�E����h���A���FCytochrome c oxidase�����ǁi�����d�nj^�j�̗Տ��I�����@���{�����Ȋw��G���@91��5���A1224-1230�ŁA1987�N�D
�@�E�R���ρF�V���ƔM�Y���@�\�ቺ�@���{�Ύ��V��@No.4255�i2005�N11��12���j�A30-32�D
�@�E�s��a�v�F�זE�ɂ�����b��B�z�������̍�p�@���@�M�B�㎏�@35(6): 707-716�A1987�N�D
�@�E�F�ŕς��@���ŕς��@�앨�̐���A����_�ƁA148-157�A2004�N9�����A�_�R������������D
�@�EGerhard Gottschalk: Bacterial Metabolism�F���c�����A��ؗ�q�@��A�P�{���_�@�Ė�F�ۂ̑�ӁA�ߑ�o�Łi1980�N�j�D
�@�EAdam Szewczyk, Lech Wojtczak(2002) Mitochondria as a Pharmacological Target. Pharmacological Reviews 54:101-127.
�@�EBernardi P (1992) Modulation of the mitochondrial cyclosporin A-sensitive permeability transition pore by the proton electrochemical gradient. Evidence that the pore can be opened by membrane depolarization. J Biol Chem 267: 8834-8839.
�@�ETrost LC, Lemasters JJ.: Role of the mitochondrial permeability transition in salicylate toxicity to cultured rat hepatocytes: implications for the pathogenesis of Reye's syndrome. Toxicol Appl Pharmacol. 1997 Dec;147(2):431-41.
�@�EVane JR, Bakhle YS, Botting RM. Cyclooxygenases 1 and 2. Ann Rev Pharmacol Toxicol 1998;38:97-120.
�@�b�g�b�v�y�[�W�b�����ƌ����̊W�b�~�j��w�m���b�����w�̒m���b��w�̘b���b�����Ȏ����b�����̕s�v�c�b�����N�W�b