�@
�@ �@
�@ �@
�@
�@�@�@
�@Th1�זE��Th2�זE
�@�����p���ɂ́AT�זE�ƁA�R�́i�Ɖu�O���u�����j���Y������B�זE�Ƃ�����B
�@T�זE�ɂ́A����ɁA�P���E�}�N���t�@�[�W����R�������A�Ɖu�����߂���A�w���p�[�s�זE�iCD4�R���z���j�ƁA�E�C���X�����זE�Ȃǂ����Q����A�L���[T�זE�iCD8�R���z���j������B
�@�w���p�[�s�זE�ɂ́ATh1�זE��Th2�זE�Ƃ�����B
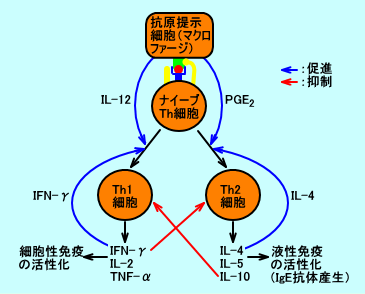
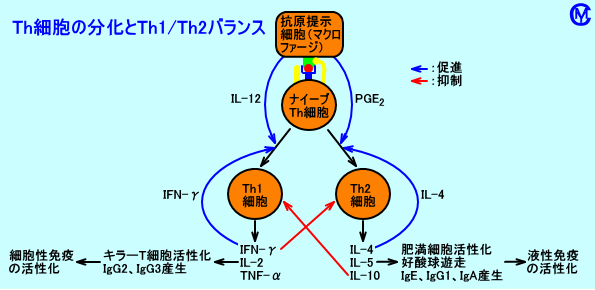
�@�R���זE���AIL-12���Y�����邩�A����Ƃ��APGE2���Y�����邩���ATh1�זE�i�זE���Ɖu�j�ƁATh2�זE�i�t���Ɖu�j�̂ǂ��炪�D�ʂɂȂ�̂��A���肵�Ă���B
�@���m�[���_�iLA�j���A���L�h���_�̐ێ�ʂ������ƁAPGE2�̎Y�����ߏ�ɍs���ATh1�זE�ɂ��זE���Ɖu���ቺ���A���M���Ԃ��������i�M�����������j�ATh2�זE�ɂ��R�̎Y�����ߏ�ɍs���A�A�����M�[�̎��ɂȂ�₷���Ȃ�ƁA�l������B
�@�P�D�w���p�[T�זE�ɂ��Ɖu�����̒���
�@�w���p�[T�זE�iCD4�z���זE�j�̑啔���́A������T�C�g�J�C���Y���p�^�[����L���Ȃ��A�i�C�[�u�w���p�[T�זE�i�i�C�[�uTh�זE�FTh0�זE�j�B
�@Th0�זE�i�i�C�[�uTh�זE�j�́A�R���זE�i�}�N���t�@�[�W�A����זE�Ȃǁj���Y������T�C�g�J�C���ɂ��A�זE���Ɖu�Ɋ֗^����Th1�זE���A�t���Ɖu�Ɋ֗^����Th2�זE�ւƁA��������B
�@���j�DTh1�זE�iT helper 1 cell�j
�@�R���זE�ł���}�N���t�@�[�W���A�R����T�זE�ɒ���ۂɕ��傷��IL-12�́ATh0�זE�i�i�C�[�uTh�זE�j���ATh1�זE�iT helper 1 cell�j�ɕ���������B
�@Th1�זE�́AIL-2�AIFN-���iIgE�R�̂̎Y����}������j�ATNF-���ATNF-���AGM-CSF�AIL-3�i���P�j���Y�����AT�זE��A�P���Ȃ��ÐH�זE�̊��������߁A�זE���Ɖu�i�c�x���N���������Ȃǁj�Ɋ֗^����B
�@Th1�זE���Y������IFN-���́ATh0�זE�i�i�C�[�uTh�זE�j��Th1�זE�ւ̕������A���i������B
�@���j�DTh2�זE�iT helper 2 cell�j
�@�}�N���t�@�[�W���A�R����T�זE�ɒ���ۂɕ��傷��PGE2�́ATh0�זE�i�i�C�[�uTh�זE�j���ATh2�זE�iT helper 2 cell�j�זE�ɕ���������i���Q�j�B
�@Th2�זE�́AIL-3�AIL-4�iIgE�R�̂̎Y����������T�C�g�J�C���ŁA�얞�זE��ANKT�זE������Y�������j�A�h�k-5�AIL-6�AIL-10�AIL-13���Y�����A�t���Ɖu�i�R�̎Y���j�Ɋ֗^����BIL-10�́ATh1�זE�����IFN-���̎Y���AIL-12�̎Y����}������B
�@Th2�זE���Y������IL-4��AIL-6�́ATh0�זE�i�i�C�[�uTh�זE�j��Th2�זE�ւ̕������A���i������B
�@Th0�זE�i�i�C�[�uTh�זE�j���ATh2�זE�ɕ�������ɂ́AIL-4���A�A���L�h���_���琶�������PGE2�̕����A�d�v�ƍl�����Ă���B
�@Th2�זE�́A�R���זE�Ƃ��āAB�זE�iIL-12���Y�����Ȃ��j����R���h�����Ă��A���B����B
�@�Q�DTh1�זE��Th2�זE�̖Ɖu�����̈Ⴂ
�@Th1�זE�ɂ���Ɖu�����ł́A�זE���Ɖu�������āA�����p����}�N���t�@�[�W�ȂǒP�j�זE���S�̉��ǔ������N��B�^�ۂ̃N���v�g�R�b�J�X�ɑ���Ɖu�����ł́ATh1�זE���D�ʂɓ����ƁA���łȓ����`������A�������Ǐ��ɕ������߂���B
�@�������ATh2�זE���D�ʂɓ����ƁA�������זE�Z�����ɂ߂ĖR�����B�Ⴆ�A�t���Ɖu�ł́A�N���v�g�R�b�J�X�Ȃǂ̍זE���ۂ��A�E���Ȃ��B���ׁ̈A�x�E�o�ɃN���v�g�R�b�J�X���[�����āA�������A�e�ՂɌ��s���ɍL�����āA�������Ȃǂǂ���B
�@Th1�זE���ATh2�זE���D�ʂɓ����Ă����Ԃł́AIgE�R�̎Y�����������A�A�����M�[�̎��Ɋׂ�₷���ƍl�����Ă���B
�@�ۑ̐����iPathogen-associated molecular pattern�FPAMP�j�́A����זE�ɍ�p���ATh0�זE�i�i�C�[�uTh�זE�j��Th1�זE�ւ̕����𑣐i���ATh1�זE�D�ʂ̏�Ԃɂ��A�A�����M�[�̎������P����B�[���A���[�O���g�A�Ȃǂ̐H�ނ�ێ悷��ƁA�A�����M�[�̎������P�����ƌ����iHygiene hypothesis�j�B
�@�E�C���X�����ł́A1�^�C���^�[�t�F�����iIFN-����IFN-���j���Y�������B1�^�C���^�[�t�F�����iType-I IFN�j�́A�s�זE�ɍ�p���AIFN-����IL-10���Y��������B
�@�ۊ����ł́A2�^�C���^�[�t�F�����iType-II IFN�j��IFN-�����Y������ATh1�זE���U�������B
�@�זE�����ہi���j���A�T�����l���ہA���X�e���A�ۂȂǁj�ɂ�銴���ǂł́A��ɁATh1�זE���U������ATh1�זE����Y�������IFN-���ɂ��H�זE�i�}�N���t�@�[�W�j������������A�܂��ATh1�זE����Y�������IL-2�ɂ��CD8�z���L���[T�זE������������A�E�ۓ����s����B
�@�זE�O�ő��B����ہiStaphylococcus�Ȃǂ��O�����z�����ۂȂǁj�ɂ�銴���ǂł́A��ɁATh2�זE���U������ATh2�זE����Y�������T�C�g�J�C���ɂ��A�R�̂��Y������A�E�ۓ����s����B
�@���̂̍R�_���\���ቺ����i�R�_����������������j�ƁATh1�זE���ATh2�זE���D�ʂɓ����ƍl������B
�@�r�^�~��C�i90mg/���j�A�r�^�~��E�i20mg/���j�A��-�J���e���i0.75mg/���j�Ȃǂ̍R�_�������́ATh2�זE�D�ʂ̏�Ԃ����P����ƍl������B
�@�A���L�h���_�i������A���ނɊ܂܂��j�̉ߗʐێ�́APGE2�̐����𑣐i�����ATh2�זE�זE�D�ʂɂ��āA�A�����M�[�̎��ɂ���̂����m��Ȃ��B
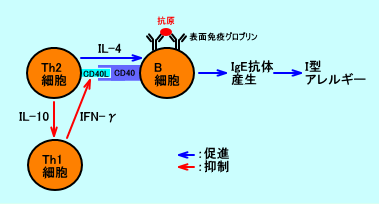
�@�R�DTh1�זE��Th2�זE�́A���݂ɗ}��������
�@Th1�זE�́AIL-2���Y�����A�L���[T�זE��NK�זE�Ȃǂ������������A�זE���Ɖu��������������B
�@Th2�זE�́AIL-4���Y�����ACD40���K���h�iCD40L�Agp39�j����āAB�זE�������������A�h�^�A�����M�[�������N����IgE�R�̂̎Y���𑣐i�����A�t���Ɖu��������������B
�@Th1�זE�́AIFN-�����Y�����邪�AIFN-���́ATh2�זE��CD40���K���h�iCD40L�j������}�����AIgE�R�̎Y����}������B
�@�܂��ATh2�זE�̎Y������IL-4��AIL-10�́ATh1�זE�̔�����}������B�@
�@IL-4��IL-10�̎Y��������ȏł́A�זE���Ɖu���}������A���X�e���A��ᚋہi�炢�ہF�n���Z���a�̌����ہj�̊������A�d�lj�����B
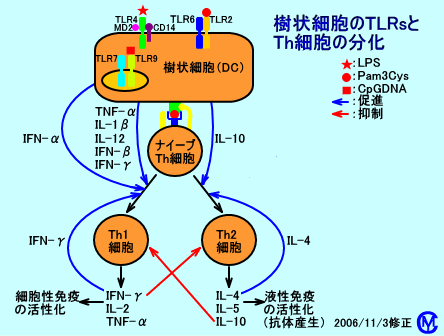
�@�S�D�R���זE��TLRs�ɂ��Ɖu�����̑I��
�@�R���זE�ł���A�}�N���t�@�[�W�iM�Ӂj�����זE�\�ʂɂ́AToll-like receptor �iTLRs�F�g���l��e�́j�����݂���BToll receptor�iToll��e�́j�́A�V���E�W���E�o�G�ɑ��݂��A�^�ۂ̊����h��Ɋ֗^����FToll��e�̂́A�n�G�i���j�̈�`�q�ŁA�^�ۂ̐N�����@�m���āA�������V�O�i����`�B���A�����h�䔽����U������B
�@�R���זE�ɑ��݂���TLRs�iToll-like receptor �j�́A�ہA�^�ہA�E�C���X�A���ȂǁA������a���̂̍\�������i�a���̂ɓ��ٓI�Ȓ`���A�őf�ADNA�ARNA�Ȃǁj��F������B�R���זE�́ATLRs�ɂ��A�a���̂̍\��������F���i���m�j����ƁA�זE���V�O�i���`�B�o�H������������A���ǐ��T�C�g�J�C����A�C���^�[�t�F�������A�Y�������B
�@�R���זE�i����זE�j�\�ʂ�TLR1�iTLR2�Ƌ����I�ɓ����j���A�ۃ��|�y�v�`�h���y�v�`�h�O���J����F���i���m�j����ƁA�R���זE���ŁANF-��B����āA�������V�O�i�����`�B����A���ǐ��T�C�g�J�C���iIL-12��AIL-6��ATNF-���j���A�Y��������BTLR1���h�������ƁA�R���זE����AIL-12���Y������ATh1�זE���������A�זE���Ɖu���U�������B
�@�܂��A�R���זE�i����זE�j�\�ʂ�TLR3�ATLR7�ATLR9���A�E�C���X�R����dsRNA�AssRNA�ACpGDNA��F������ƁA�R���זE���ŁA�������V�O�i�����`�B����AI�^�C���^�[�t�F�����iIFN-���AIFN-���j���A�Y��������iTLR3�́AIFN-���݂̂��Y��������j�BTLR4�́A�ۗR���̃��|�|���T�b�J���C�h�iLPS�j��A�E�C���X�̒`���iRS�E�C���X�̗Z���`���j��F�����AIFN-�����Y����������A���ǐ��T�C�g�J�C�����Y��������B
�@���̂悤�ɁA�R���זE�i����זE�j�́A�\�ʂ�TLRs�ɂ��A���̂ɐN�������̂��A�a���̂Ȃ̂��i�ۂȂ̂��A�E�C���X�Ȃ̂��j�A�őf�Ȃ̂��Ȃǂ����m���A����ɉ����āA���ǐ��T�C�g�J�C�����Y��������A�C���^�[�t�F�������Y������B���̌��ʁATh0�זE�i�i�C�[�uTh�זE�j�́ATh1�זE�i�זE���Ɖu�F�a���̂̋ۑ̂��ÐH����j��ATh2�זE�i�t���Ɖu�F�a���̂̓őf�𒆘a����j�ɕ�����������B�@���n����זE�iiDC�j�\�ʂ�TLRs���A�ۗR���i�咰�ۂȂǂ̋ۑ̐����j��LPS�i���|�����j��CpG�I���S�k�N���I�`�h�iCpGDNA�j�A�E�C���X�R���̓�d��RNA�idsRNA�j��F�����A�זE���Ɏ�荞�ނƁA���n����זE�iiDC�j�́AIFN-�����Y������DC1�֕������ADC1�́A��荞�y�v�`�h�R���iLPS�Ȃǁj���AMHC�N���XII���q�Ƌ��ɁA�זE�\�ʂɔ�������B����DC1�\�ʂ̕����́i�y�v�`�h�R���{MHC�N���XII���q�j���A�����p�߂�Th0�זE�i�i�C�[�uTh�זE�j���AT�זE�R����e�́iTCR�j�ŔF�����ADC1���Y�������IFN-���ɂ��h�������ƁATh1�זE�֕������AIFN-�����Y������ƌ����B
�@���n����זE�iiDC�j�\�ʂ�TLRs���A���R���̍R����A�J�r�R���̍R����F�����A�����������ƁADC2�֕�������BDC2�́AIL-4���Y�����ATh0�זE�i�i�C�[�uTh�זE�j���ATh2�זE�֕�������ƌ����B
�@����זE�iDC�j�����IL-12����i�Y���j�́AIFN-���ACD40L�A�ہi���F�u�h�E���ۂȂǁj��ۗR���̕����iLPS�ACpG�j�A���i�g�L�\�v���Y�}�Ȃǁj�ɂ���đ��i�����B
�@����זE�iDC�j�����IL-12����i�Y���j�́APGE2�A�T�C�g�J�C���iIL-10 �AGM-CSF�j�A�P���J�C���iMCP-1�Ȃǁj�A��́iC5a�j�ACD47�ithrombospondin receptor�j�A�X�e���C�h�z�������iCorticosteroids�j�A�E�C���X�i���]�E�C���X�j�Ȃǂɂ��A�}�������B
�@TLR4��LPS�Ŏh��������ATLR9��CpG DNA�i�ۂ�DNA�j�Ŏh������ƁA����זE�����n���A����זE����IL-12�iIL-12p70�j�����o����ATh1�זE���D�ʂɗU�������B�������A������iIFN-�����݉��j�ł́A��ʂ̋z�����ꂽLPS�́ATh2�זE��U�����A�A�����M�[�����������N�����BTLR4��LPS����������ƁATRIF��TRAM�����N���[�g����AIFN-�����Y�������BCpG�́ATh1�זE���֗^����IgG2a�T�u�N���X�R�̂̎Y����������BTh1�זE���֗^����IgG2a�T�u�N���X�R�̂̎Y���ɂ́A�������ɎY�������I�^�C���^�[�t�F�����iIFN-����IFN-���j�̑��݂��K�{�BAlum�ialuminum hydroxide gel�j�́ATh2�זE���֗^����IgG1�T�u�N���X�R�̂̎Y����������iEnrico Proietti���j�B
�@TLR3�ATLR4�ATLR7�ATLR9�����IFN-���h���ɂ��A����זE����IL-12�iIL-12p70�j�����o����ATh1�זE���U�������B
�@TLR5�ATLR9�A���邢�́ATLR3�́AIL-12�A���邢�́AIFN-���̕��o����āATh1�זE��U������B
�@���ɁATLR2���h�������ƁA����זE����͏��ʂ�IL-12���Y������ATh2�זE���U�������BTLR2���AV�R���iYersinia enterocolitica V antigen�j��z�X�t�@�`�W���Z�����iphosphatidylserine from Schistosoma mansoni �j�ɂ��h�������ƁAIL-10�����o����ATh2�זE���U�������BPam3Cys�i�ۂ̃��|�y�v�`�h�j�́ATLR2�Ɍ������ATh2�זE��U������B
�@�R�E�C���X���Imiquimod�́ATLR7��TLR8���h�����AIFN-�����Y��������B
�@�A���z�e���V��B�iamphotericin B�FStreptomyces nodosusM4575���R���j�́ATLR2�Ɍ�������ׁA����p�Ƃ��Ĕ��M��������B
�@����זE�iDendritic cells�FDC�j�́A���t���A�����p�g�D���Ȃǂɑ��݂���B
�@����זE�́ACD4�z���w���p�[T�זE�ɁA�O���R���̍R�������B
�@����זE�ɂ́A�~�G���C�h�n�i�����n�j����זE��myeloid DC �iMDC�j�ƁA�����p���n����זE��plasmacytoid DC �iPDC�j�́A��̈قȂ�W�c�ipopulations�j�����݂���B
�@myeloid DC �iMDC�j�́A�\�ʃ}�[�J�[�́ACD11c+�ACD123dim�AHLA-DR+�ŁA�P���R���̎���זE�Ɠ����Bmyeloid DC �iMDC�j�́A�P���w���y�X�E�C���X�iHSV�j�h���ɂ���ẮAIFN-�������܂�Y�����Ȃ��B
�@plasmacytoid DC �iPDC�j�́A�\�ʃ}�[�J�[�́ACD11c-�ACD123bright�AHLA-DR+�ABDCA-2+�ABDCA-4+�Bplasmacytoid DC �iPDC�j�́AIFN-�����ʂɎY������B
�@myeloid DC �iMDC�j�́Apre-DC1�זE�ł���A�������P���R������זE���AM-CSF�AIL-4�ACD40-ligand�iCD40L�j�h���ɂĔ|�{����ƁAIL-12���Y������ADC1�זE�ɐ��n���A�i�C�[�uT�זE��Th1�זE�iIFN-�����Y������j�ɕ���������B
�@lasmacytoid DC �iPDC�j�́Apre-DC2�זE�ł���AIL-3�ɂĔ|�{����ƁADC2�זE�ɐ��n���A�i�C�[�uT�זE��Th2�זE�iIL-4���Y������j�ɕ���������B
�@DC2�זE�ɂ���ėU�����ꂽTh2�זE���Y������IL-4�́ADC2�זE��}�����邪�ADC1�זE���͑��i����B
�@DC1�זE�ɂ���ėU�����ꂽTh1�זE���Y������IFN-���́ADC2�זE�ɂ���ėU�����ꂽTh2�זE���Y������IL-4�iDC2�זE��}������j�̍�p��}���i���a�j����B
�@myeloid DC �iMDC�j�́AIL-12���Y�����邪�A�T�^�C���^�[�t�F�����iIFN-���j�����܂�Y�����Ȃ��Bplasmacytoid DC �iPDC�j�́A�T�^�C���^�[�t�F�����iIFN-���AIFN-���j���Y�����邪IL-12���قƂ�ǎY�����Ȃ��B
�@plasmacytoid DC �ipre-DC2�FTh2�זE��U������j�́ACD40-ligand�iCD40L�j�h���ɂ��A�T�^�C���^�[�t�F�����ɉ����Amyeloid DC �ipre-DC1�j�Ɠ��l��IL-12���Y������iTh1�זE��U������j�Ƃ�������B
�@myeloid DC �ipre-DC1�j�́AIL-10�ŏ�������ƁAIL-12�̎Y�����}������ATh2�זE��U������B
�@PGE2��A�X�e���C�h�z���������AIL-10�Ɠ��l�ɁA�imyeloid DC����́jIL-12�̎Y����}������B
�@�q�g�ł́A�V���ɕ������ꂽ�i�h�����Ă��Ȃ��jmyeloid DC�́ATLR1�ATLR2�ATLR3�ATLR5�ATLR6�ATLR8�����Ă���B�����Aplasmacytoid DC�́ATLR7�ATLR9�����Ă���B�}�E�X�ł́A���ҁimyeloid DC��plasmacytoid DC�j���ɁATLR1�ATLR2�ATLR4�ATLR6�ATLR8�ATLR9�����ATLR3�́A myeloid DC�݂̂��������Ă���B
�@plasmacytoid DC�i�`���זE�l����זE�j�́A�T�^�C���^�[�t�F�����iIFN-���j�̎Y���\�������B
�@TLR7�ATLR9����ẮAINF-���̎Y�����U�������Bplasmacytoid DC�i�`���זE�l����זE�j�́ATLR7�ATLR9�����Ă��āAIFN-���̎Y���\�������B
�@TLR3�ATLR4����ẮAINF-���̎Y�����U�������B
�@TLR2����ẮiTLR2�h���ł́j�AI�^�C���^�[�t�F�����iIFN-����IFN-���j�̎Y���́A�U������Ȃ��B
�@����זE�́A�C���^�[�t�F����-���iIFN-��2a�j�̑��݉��ACD40L�ɂ���Ďh�������ƁAIL-10�iTh2�זE��U������j��IL-12�iTh1�זE��U������j�̎Y�����A��������B
�@����זE�́A�C���^�[�t�F����-���iIFN-��2a�j�̑��݉��ALPS�ilipopolysaccharide�j�ɂ���Ďh�������ƁA��ɁAIL-10���̂ݎY������B
�@�C���^�[�t�F�������Y������זE�iIPCs�j�́ACD4+CD11c-2�^����זE�O��זE�ipDC2s�j�ł���B
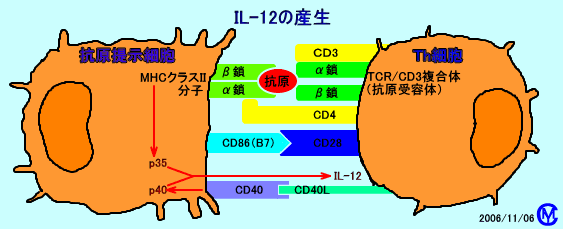
�@IL-12�́Ap40��p35�ɂ���č\�������i�w�e���_�C�}�[�j
�@�R���זE����R���i�R���h���j����Th�זE���A�זE�\�ʂ�CD40-ligand�iCD40L�FCD154�j�����A�R���זE�\�ʂ�CD40���h������iCD40/CD40L���h���o�H�j�ƁA�R���זE�i����זE�Ȃǁj������������AIL-12p40������B
�@�R���זE����R������Th�זE���AT�זE�R����e�́iTCR�j�ɂ��A�R���זE��MHC�N���XII���q���h������ƁA�R���זE��IL-12p35������B
�@IL-12p40��IL-12p35���A�w�e���_�C�}�[���`�����A����������L����IL-12���Y�������B
�@�������ɁA��q�����Ŏ���זE���A�����Q���ƃG���h�g�L�V���֓����\�I�����ƃ������[Th1�זE��}����T�זE���D�ʂƂȂ�A����זE���A�����Q���ɂ̂ݖ\�I�����ƃ������[Th2�זE���D�ʂɂȂ�B
�@�G���h�g�L�V���̗ʂ́A�z���R���ɑ���Th2�זE�̕����U���ɁA�e�����y�ڂ��B
�@���n����זE�iIL-2�Y���\���Ⴂ�j���A�R���Ɠ����ɁA��ʂ̃G���h�g�L�V����TLR4���h������ƁATh2�זE�ւ̕������U�������B�����A���ʂ̃G���h�g�L�V����TLR4���h������ƁA����זE��IL-2�Y���\����������ATh1�זE�ւ̕������U������Ղ��Ȃ�ATh2�זE�ւ̕������}�������B
�@����זE��TLR2���y�v�`�h�O���J���Ŏh������ƁA��ɁAIL-6�AIL-10���Y������邪�AIL-12�̎Y���͏��Ȃ��B
�@����זE��TLR3���{��RNA�Ŏh������ƁAIL-6�AIL-10�̎Y���͏��Ȃ����AIL-12�̎Y���͑����B
�@����זE���G���h�g�L�V���Ŏh������ƁAIL-6�AIL-10�AIL-12���ɁA�Y�������B
�@�T�DNKT�זE
�@NKT�זE�́AIFN-�����AIL-4�����A�Y������B
�@IFN-���AIL-4�́A���������p�����F�h�e�m-���́ATh1�זE�������������AIL-4�́ATh2�זE��������������B
�@�ENKT�זE�́A�R���זE�i����זE�j��CD1�����q�iMHC�N���X�hb���q�j�Ɍ��������A�X�t�B���S�������i��-�K���N�g�V���Z���~�h�F��-GalCer�j���A�R����e���ŔF������ƁA����������AIL-4�Ƃh�e�m-�����Y������B
�@�E�X�t�B���S�������i��-�K���N�g�V���Z���~�h�F��-GalCer�j�́A����זE�i�R���זE�j��IL-12���Y��������B�R���זE����Y�����ꂽIL-12�́ANKT�זE�ɍ�p���A�h�e�m-�����Y����������AFas���K���h����āA�זE��Q��������������B�@
�@�ENKT�זE�́A��1,3-�I���S���ɂ��A�זE�\�ʂ�CD161���h�����ꊈ���������ƁA�h�e�m-���݂̂��Y������B
�@�ENKT�זE�́A�R����e�̂���Ďh�������ƁAIL-4���Y������B
�@�ENKT�זE�́A�X�t�B���S�������̗U���́iOCH�Ƃ��������������j�ɂ��h�������ƁAIL-4�݂̂��Y������Ƃ����B
�@�U�DIL-18
�@IL-18�́A�A�����M�[�������̔��ǁiIgE�Y����A�����M�[�����ǁj�Ɋ֗^����B
�@IL-18�́A�}�N���t�@�[�W���őO��̂Ƃ��ĎY������A�O�����A���ۂ̍זE�ǂ̃��|�����́iLPS�j�ɂ�芈�������ꂽcaspase-1�ɂ��ؒf����A�����^�ƂȂ�A���傳���B
�@IL-18�́ATh1�זE���A�R���Ŏh�����i�R�����݉��Ɂj�A�����������A�h�e�m-�����Y��������B�܂��AIL-18�́A�iIL-2����������ƁA�jTh2�זE���A�����������ATh2�T�C�g�J�C���iIL-4�AIL-5�AIL-9�AIL-13�j���Y��������B
�@IL-18�́A�}�E�X�ɓ��^����ƁA�R���h�������ɁAT�זE�������������AT�זE��IL-4�Y���ACD40���K���h�iCD40L�j�����������AIgE�Y����U������B
�@IL-18�́AIL-3�Ƒ���I�ɁA�얞�זE�i�}�X�g�זE�j��A�D����ɍ�p���āAIL-4�AIL-13�A�q�X�^�~���Y����U������B
�@�����ǂ́A�A�����M�[�������̏Ǐ���A���������邪�A�a���̐����́A�C�����זE��A�畆�P���`�m�T�C�g���h�����AIL-18���Y��������B
�@�}�E�X���A�g�s�[���畆�����f���ł���NC/Na�}�E�X�iNC�}�E�X�j�́A���ۉ��Ŏ��炷��ƁA�A�g�s�[���畆���ǂ��Ȃ����A�ʏ�̊����Ŏ��炷��ƁA�q�g�̃A�g�s�[���畆���Ɠ��l�Ȕ畆���ǂ���B
�@�����p���́A�X�[�p�[�R���ł���SEB��A�O�����A���G���h�g�L�V���ł���LPS�Ŏh�������ƁA�h�e�m-�����Y������BNC�}�E�X�̃����p���́A�X�[�p�[�R���iSEB�j��A�O�����A���G���h�g�L�V���iLPS�j�Ŏh�������ۂ̂h�e�m-���Y���iTh1�T�C�g�J�C���Y���j���A�ቺ���Ă���BNC�}�E�X�̃����p���́AV��8T�זE��e�́i�R����e�̂̃����j�̈�`�q���������AV��8+T�זE��AV��8+NKT�זE���A���@���Ă���ׁASEB�ɂ��h�e�m-���Y�����A�ቺ����B�܂��ANC�}�E�X�̃����p���́A�h�e�m-���Y���U�����q�ł���IL-18�́A�}�N���t�@�[�W����̎Y�����ቺ���Ă���ׁALPS�ɂ��h�e�m-���Y�����A�ቺ����B
�@NC�}�E�X�̃����p���́A�RCD3�R�̂Ŏh�������ہAIL-4�Y���iTh2�T�C�g�J�C���Y���j���������Ă���B
�@���̂悤�ɁANC�}�E�X�́A��ۂ̎h���ɂ��h�e�m-���Y���iTh1�T�C�g�J�C���Y���j���A�ቺ���A�܂��AIL-4�Y���iTh2�T�C�g�J�C���Y���j���A�������Ă���ׁATh2�זE���D�ʂɕ������AIgE�R�̎Y�����������A�A�g�s�[���畆���Ɠ��l�Ȕ畆�������ǂ���ƁA�l�����Ă���B
�@NC�}�E�X���A�h�e�m-���AIL-12�A���́AIL-18�Ŏ��Â���ƁA����IgE��IL-4���ቺ���A�畆�������P����B
�@
�@�V�D�זE���Ɖu�Ɖt���Ɖu
�@�זE���Ɖu�ł́ATh1�זE���Y������IFN-���ɂ��A�}�N���t�@�[�W������������A�זE���ۂ��A�E�ۂ����B�܂��ATh1�זE���Y������IL-2�ɂ��A�L���[T�זE������������A�E�C���X�����זE���A��Q�����B
�@�t���Ɖu�ł́ATh2�זE���Y������IL-4�AIL-5�AIL-6�AIL-13�ɂ��AB�זE�������E���B���A�R�́i�Ɖu�O���u�����j���A�Y�������B�R�̂́A�a���̂��Y������ۑ̊O�őf�𒆘a���A�זE�O�ۂ��I�v�\�j�������}�N���t�@�[�W�Ȃǂɂ���ÐH�𑣐i���A��̌n�����������n�ۂ���B
�@�W�D�����ۂ�Th1/Th2�o�����X
�@�V�����̖Ɖu�n�́ATh2�זE���A�D�ʂɓ����Ă���B
�@�Ɖu�n��Th1/Th2�o�����X�iTh1�זE��Th2�זE�̔䗦�j�́A�����ǂ��������p�ɂ���āA�e������B���ۃ}�E�X�ł́A������������ATh2�זE���A�D�ʂɓ����Ă���B
�@��������ɁA�R�������𓊗^���A�����ۑp����������i�ω�������j�ƁA�������Ă�����ATh2�זE���A�D�ʂɓ����Ă���i�A�����M�[�������ɂȂ�Ղ��j�B2�܂łɍR�������̓��^������A�S���P���N�`���ڎ������A��e���A�g�s�[�������̊�����L���Ă���ƁA�A�g�s�[�������ǂ��Ղ��B
�@2�̏����̕��ւ������������ʂł́A�A�����M�[�늳���ł�Staphylococcus aureus�i���F�u�h�E���ہF�D�C���ہj���L�ӂɑ����ALactobacilli��Bifidobacteria�����Ȃ������ƕ���Ă���B
�@�X�DRS�E�C���X������Th1/Th2�o�����X�@
�@RS�E�C���X�iRSV�j�́A��������ƁA�S�`�U���̐������Ԃ̌�ɁA�C�ǎx���ǂ���B
�@RS�E�C���X�����ǁi�C�ǎx���Ȃǁj�ł́A�b�a����B
�@RS�E�C���X�́A�����O����L���Ă��āAG�iattachment�j�AF�ifusion�j�ASH�ismall hydrophobic�j�̂R��ނ̕\�ʒ`�������݂���B
�@G�`���́ARS�E�C���X���h��זE�\�ʂڒ�����̂ɕK�v�ŁAF�`���́ARS�E�C���X���h��זE���N������̂ɕK�v�BG�`���́ATh2�T�C�g�J�C���iIL-4�AIL-5�j���Y�������A�D�_����V��������i�D�_���Z�����N�����j���AF�`���́ATh1�T�C�g�J�C���iIFN-���j���Y�������A�P�j���Z�����N�����i�L���[T�זE���������E�U������j�B
�@RS�E�C���X�ɑ���L���[T�זE�i�זE��Q��T�����p���j���A�Ɖu�}���}�E�X�ɒ�������ƁA���ʂ̏ꍇ�͋C���a�ς𗈂�������RS�E�C���X�̑��B���}������邪�A���ʂ̏ꍇ�͏o�����x�������N����iTh1�זE�ɂ��ARS�E�C���X�ɑ���L���[T�זE���ߏ�ɗU�������ƁARS�E�C���X�����ǂ��d�lj����邨���ꂪ����j�B
�@1960�N��ɕč��Ńz���}�����s����RSV���N�`���̐ڎ킪���݂�ꂽ���A�z���}�����s����RSV���N�`���́ARS�E�C���X�����ǂ̗\�h���ʂ��Ȃ���A�ڎ��������RS�E�C���X�Ɏ��R��������ƁA�d�lj����邱�Ƃ�����B
�@�z���}�����s����RSV���N�`����ڎ��ɁARS�E�C���X�Ɏ��R���������S���������́A�x�ɍD������D�_���������ɐZ�����Ă����i�ʏ�ARS�E�C���X�ɂ��C�ǎx���ł́A�x�ɍD�_����D�����̐Z���͔F�߂��Ȃ��j�B�z���}�����s����RSV���N�`���́AG�`������āATh2�זE��U�����iTh2�������N�����j�A�D�_����D�����̒����ȐZ�����N�����i�D�_���̐Z���́ATh2�זE�ɂ��Th2�T�C�g�J�C���̎Y����A�얞�זE�Ȃǂ�������C�R�g���G���̎Y������������j�B
�@RS�E�C���X�ɂ��C�ǎx���ł́A�@��o�t���Ɋ܂܂��Th2�T�C�g�J�C���iIL-4�AIL-5�AIL-13�j�͌��o���x�ȉ��ƌ�����i�C�ǎx���̔��ǂɂ́ATh2�זE�ɂ��R�̎Y���̊֗^�͏��Ȃ��ƍl������j�B�C�ǎx���┽�����b���̎��ł́A�@�����t����IFN-��/IL-4��́A��C�����̎��ɔ䂵�āA�L�ӂɍ��l�������B�]���āA�iRS�E�C���X�ɂ��j�C�ǎx���ł́A�iTh2�זE���D�ʂȋC�ǎx�j�b���ƈقȂ�ATh1�זE���D�ʂɗU������Ă���ƍl������iRS�E�C���X�ɂ��C�ǎx���ł́ATh1�זE�ɂ��A�L���[T�זE���U������A�b�𗈂����j�B
�@�V�i�W�X�iSynagis�FPalivizumab�F�p���r�Y�}�u�j�́A�RRS�E�C���X�q�g�����m�N���[�i���R�́i�}�E�X�RRS�E�C���X���m�N���[�i���R�̂̑�����蕔�ʁ{�q�gIgG1��핔�y�щϕ��t���[���z��j�B
�@�V�i�W�X�́ARS�E�C���X��F�`���i�R������A�̈�j�ɑ�����ٓI�q�g�����m�N���[�i���R�́B�V�i�W�X�́AF�`���iRS�E�C���X���h��זE�ɐڒ��E�N������ۂɏd�v�Ȗ������ʂ����j�Ɍ������āA�E�C���X�̊������𒆘a���A�E�C���X�̕����y�ё��B��}������B
�@�V�i�W�X�́A�K��������V�����A�����A�c���i24�J����ȉ��̐�V���S���������Ȃǁj�ɑ��āARS�E�C���X�̊������s�����ɁA�d�Ăȉ��C�������̔��Ǘ}����ړI�Ƃ��āA�̏d����P�Tmg�i15mg/kg�j���A��1��A�ؓ����˂���i���˗ʂ�1mL���z����ꍇ�ɂ́A�������^����j�B
�@�V�i�W�X�́ARS�E�C���X�ɓ��ٓI�ɍ�p����̂ŁA�\�h�ڎ�i���N�`���ڎ�j�ɂ��Ɖu������W���Ȃ��ƍl�����Ă���i�V�i�W�X���^�������́A�O�퍬���\�h�ڎ��ABCG�\�h�ڎ�Ȃǂ��Ă��\��Ȃ��j�B
�@RS�E�C���X�ɂ��C�ǎx���̊����i2�Ζ����j�ɁA�f�L�T���T�]���iDEX�j��P��0.4mg/kg�牺���˂���ƁA�R���Ǎ�p�ɂ�艺�C���Ǐ�̑������}������A���@�����ቺ����ƌ����B
�@�P�O�D�T���R�C�h�[�V�X��L�^�A�N�l�ەa������Th1�זE
�@�T���C�R�h�[�V�X�isarcoidosis�j�́A�S�g������i�x�A�����p�߁A�畆�A���A�̑��A�B���A�S���A���i�A�t���A�]�_�o�Ȃǁj�ɁA�������Ȃ��ޏ��זE�������`�����鎾���B
�@�T���R�C�h�[�V�X�̕a���Ɋւ��āA�Â�����A���ď����ł́A���j�����^���ė������A�T���R�C�h�[�V�X�̕a�ϕ��ʂ��猋�j�ۂ͔|�{����Ȃ��B
�@���{�ł́A�̖{�ԓ��b�������̓�a�����ǂɂ���āA1970�N��ɁA�a�ϕ��ʂ���A�N�l�ہiPropionibacterium acnes�j�����C�|�{�ɂĕ������ꂽ�B
�@1999�N�ɁA�T���R�C�h�[�V�X�̕a�ϕ��̃����p�߂ɂ́A��O�Ȃ��A�A�N�l��DNA��A�畆��ۂł���P. granulosum��DNA�����݂��邱�Ƃ��𖾂���A�T���R�C�h�[�V�X�̕a�����A�N�l�ۂł���\�������܂����B
�@�T���R�C�h�[�V�X�̕a�ϕ��̃����p�߁i�g�D�����t�j���}�E�X�ɖƉu���A�A�N�l�ۂɓ��ٓI�ɔ��������̃��m�N���[�i���R�̂��쐬���ꂽ�BPAB�R�̂́A�A�N�l�ۂ̃��|�^�C�R�_�i�זE���������̓������R���j��F������BTIG�R�̂́A�A�N�l�ۂ̃g���K�[�t�@�N�^�[�i���|�\�[���������̃V���x�����`���j��F������B
�@9���ȏ�̃T���R�C�h�[�V�X�̏Ǘ�ł́A�����i�̍זE�j�ɁAPAB�R�̂ŗz���������i���F�����j���^�~�`���̂��F�߂���B�T���R�C�h�[�V�X�ł́A��^�a���`���́iHW���́j�����݂���̂����������APAB�R�̂��A�i��^�a���`�j���̂̊O������芪���悤�ɁA�z���������B�܂��A���̓����́ATIG�R�̂��A�z���i�h�b�g��j�������B���̂��Ƃ���A��^�a���`���́iHW���́j�́AL�^�A�N�l�ہi�זE�Ǎ\�����������זE���ɐ����������Ă���A�N�l�ہj�ł��邱�Ƃ����������B
�@L�^�A�N�l�ۂ��A�זE�����B������ƁATh1�זE�^�Ɖu�������N����A�����^�A�N�l�ہi�~�`���^��L�^�A�N�l�ۂŁA�����p�t�⌌�t����đS�g�Ɋg�U����j�̊g�U��h�~����ׁA�����`�������i�T���R�C�h�[�V�X�̊��҂́A�Ɖu�s�S�������āA�זE���ɐ�����������L�^�A�N�l�ۂ�r���o���Ȃ��j�B
�@�R���܂��~�m�}�C�V���iMINO�j���A�N�l�ۂ̏��ۖړI�œ��^����ƁA�T���R�C�h�[�V�̊��҂̖��ɁA���ʂ��F�߂��Ă���B�~�m�}�C�V���ȊO�ɁA�A�W�X���}�C�V���A�N�����X���}�C�V���A�N�����_�}�C�V���A�o�N�^�iST���܁j�𓊗^���Ă��A�����̎��Ð����Ⴊ����Ă���B
�@�T���R�C�h�[�V�X�̗ޏ��זE�����́A�A���W�I�e���V���ϊ��y�f�iACE�j�𑽂��܂�ł���B
�@�������T���R�C�h�[�V�X���҂�80���ȏ�́AACE���l�������B
�@���t�玿�X�e���C�h���Ŏ��Â��J�n����ƁA��r�I�����ɁAACE�l���ቺ���邱�Ƃ�����B
�@�P�P�D�����ƖƉu�@�\
�@�����́A���̓��̖�300�ȏ�̍y�f�̊����Ɋ֗^���Ă��āA�����ێ��ɕK�{�Ȕ��ʌ��f�ƌ�����B
�@�����́A��ɏ\��w�������z�������B
�@����זE�́ATLR�����LPS�ɂ��h�������ƁA�זE�������Z�x���ቺ���A����������i�זE�\�ʂ�MHC�N���XII���q�̔�������������j�ACD4+T�זE�i�w���p�[�s�זE�j���U�������B
�@�זE�������Z�x�́A�Q��ނ̈����g�����X�|�[�^�[�t�@�~���[�ɂ���Ē��߂���Ă���B
�@�����A�����g�����X�|�[�^�[�t�@�~���[�ɂ́A�������זE���Ɏ�荞��ZIP�t�@�~���[�iimporter�j�ƁA�������זE�O�ɂ����o��Znt�t�@�~���[�iexporter�j�Ƃ��A���݂���B
�@LPS�ɂ�����זE�\�ʂ�TLR4���h�����A����זE��������������ƁATRIF�iMHC�N���XII���q���܂ޏ��E�̍זE���ړ��Ɋ֗^����j����āAZIP�t�@�~���[�i�������זE���Ɏ�荞�ވ����g�����X�|�[�^�[�t�@�~���[�j�̔������������A�זE�������Z�x�������ɒቺ����B
�@�����C���|�[�^�[Zip6/Liv1�́ALPS�h���i����זE��TLR4���h�������j�ɂ��ATRIF����Ĕ�������������i����זE�́A�זE�������Z�x���������A�זE�\�ʂ�MHC�N���XII���q�̔������������A�R�����ٓI��CD4+T�זE�̗U�������i�����j�B
�@�����C���|�[�^�[Zip6/Liv1��������������������זE�i�זE�������Z�x���㏸����j�́ALPS�h���ɂ��MHC�N���XII���q�����̑������}������Ă��āA�R�����ٓI��CD4+T�זE�i�w���p�[T�זE�j���U���i�h���j����Ȃ��B
�@�P�Q�D���̑�
�@�E�ۂ��N������ƁA�ۂɊ܂܂�鎉���i���b�j�ɂ���āA���Ǔ���TLR���������������B
�@���Ǔ���TLR���́A�H�����̎����i���b�j�̗ʂ������ƁA�ۂɊ܂܂�鎉���Ɓu���Ⴂ�v������������Ă��܂��B
�@���P�FIL-3�́ATh1�זE������ATh2�זE������A�Y�������B
�@���Q�F�}�N���t�@�[�W�Ȃǂ̍זE�istromal cells and tissue-infiltrating mononuclear cells�j����Y�����ꂽPGE2�́A�זE��cyclic AMP �icAMP�j�������ATh1�זE�ɂ��AIFN-����IL-2�̎Y����}������BPGE2�́ATh2�זE�ɂ��AIL-4�Y����}�����Ȃ��B�]���āAPGE2�́A�}�N���t�@�[�W����Y�������PGE2�́ATh0�זE�i�i�C�[�uTh�זE�j���ATh2�זE�iT helper 2 cell�j�זE�ɕ���������B
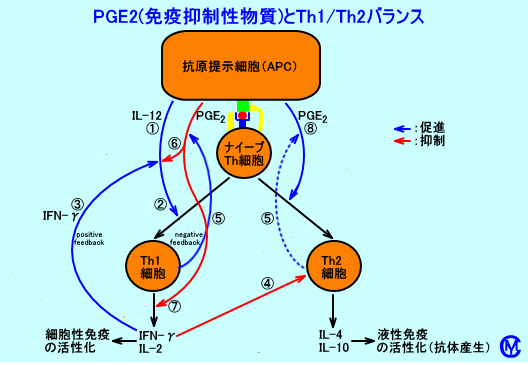
�@�@�R���זE�iAPC�j�́AT�זE�Ƃ�CD40/CD154�iCD40L�j���h���iCD40-CD154 interaction�j�ƁAIFN-���ɂ��AIL-12���Y������B
�@�A�R���זE���Y������IL-12�́ATh0�זE�i�i�C�[�uTh�זE�j���ATh1�זE�֕���������B
�@�B�R���זE���Y������IL-12�́AT�זE�iTh1�זE�j��ANK�זE����AIFN-�����Y��������BTh1�זE���Y������IFN-���́A�R���זE����IL-12���Y��������ipositive feedback�j�B
�@�CTh1�זE���Y������IFN-���́ATh2�זE�̊�������}������B
�@�DTh1�זE�́ATh2�זE��葽���A�R���זE���PGE2���Y��������FTh�זE�ɂ��PGE2�Y���́ATh1�זE��TCR���A�R���זE��MHC�N���XII���q�{�R���y�v�`�h��F�����邱�Ƃ��K�v�����ACD40/CD154���h����AIFN-���́A�K�v�łȂ��iTh2�זE���A�R���זE����APGE2���Y�������邪�ATh1�זE��菭�Ȃ��j�B
�@�EPGE2�́A�R���זE�ɍ�p���A����IL-12�Y����}�����ATh1�זE�i�ւ�Th0�זE�������j��}������idown regulation�j�B�R���זE�imicrogliaI�j�ɂ��IL-12�Y���́APGE2�ȊO�ɁAIL-10��AcAMP�Z�x���㏸��������q�icAMP-elevating agents�j�ɂ���Ă��A�}�������B�Ȃ��APGE2�́AIL-6�̎Y�����}������i�ቺ������j���APGE2�́A�iTh2�זE��}�N���t�@�[�W����Y�������jIL-10�̎Y�����́A����������i2�{���x�j�BPGE2�́ACD3�R�́immobilized anti-CD3�j�ɂ��h�����ꂽ�ACD4+ T cell clones�����. IFN-���AIL-2�AIL-4�AIL-5�AIL-10�̎Y�����́A�}������B
�@�FPGE2�́ATh1�זE�ɍ�p���ATh1�זE�ɂ��T�C�g�J�C���iIFN-����IL-2�Ȃǁj�̎Y����}�������iTh1�זE���A�R���זE�́APGE2�ɂ��Anegative feedback����j�B�Ȃ��APGE2�́ATh2�זE�ɂ��T�C�g�J�C���iIL-4�j�̎Y�����͗}�����Ȃ��B�]���āAPGE2�ɂ��ATh2�זE�̋@�\���A�D�ʂɂȂ�B
�@�GPGE2�́ATh0�זE�i�i�C�[�uTh�זE�j�ɍ�p���ATh2�זE�֕��������AIL-4�AIL-5�AIL-10�AIL-13���Y��������B�Ȃ��APGE2�́A�R���זE�����IL-12�Y���Ɉˑ������iindependant of IL-12 secretion�j�Ain the basal state�i�R���h������Ă��Ȃ���ԁj�ł́ATh0�זE�i�i�C�[�uTh�זE�j�ɍ�p���ATh1�זE�֕���������F����זE�iDC cells�j��T�זE�iCD4+ CD45RA+ T cells�j���APGE2�i10-7M�j��Y�����āA�����|�{����ƁA�|�{14���̔|�{�㐴���ɂ́A���ʂ�IFN-�������o����Ă��邪�APGE2��Y�����Ȃ��ꍇ���AIFN-���ʂ����Ȃ��B�܂��A�|�{�㐴����IL-4�ʂ́A���Ȃ��B����זE��T�זE�̔䗦�iDC/T cell ratio�j���Ⴂ�i300:1�j�ƁATh0�זE�inative T cells�j����ATh1�זE���ATh2�זE�����A�U�������B
�@���̂悤�ɁA�R���זE���Y������AIL-12��PGE2�̃o�����X�i�R���זE���AIL-12���Y�����邩�A����Ƃ��APGE2���Y�����邩�j���ATh1�זE�ƁATh2�זE�̂ǂ��炪�D�ʂɂȂ�̂��A���肵�Ă���B
�@PGE2�ɂ́A���ɂ�����i�u�����N���j�A���Ǔ��ߐ��i�����A��E�����Ȃǂ𗈂����A���M������A�ȂǁA�������i��p������B
�@�������A�}�N���t�@�[�W�i�P���j����Y�������PGE2���A�Ɖu�n�ɑ��ẮA�Ɖu������}������Ɖu�}���������iimmunosuppressant�j�Ƃ��āA���Ǘ}����p���������Ƃ������B
�@�}�N���t�@�[�W����Y�������PGE2��PGE1�́A�iin vitro�ŁA�jT�זE�̍R���ɑ��鑝�B�����A�L���[T�זE�̗U���E�����AT�זE��IL-2�Y���AB�זE�̑��B�E�R�̎Y���ANK�זE�����A�}�N���t�@�[�W�̔S�����E�L�W��}������B���̂悤��PGE2�̖Ɖu�}����p�i���Ǘ}����p�F�R���Ǎ�p�j�́A�����p���Ȃǂ�PGE2��e�́iEP2��e�́j����āA�A�f�j���_�V�N���[�[�iAC�j�������������AcAMP�������邱�ƂŁA�����B�Ȃ��APGE2��PGE1�́A�}�N���t�@�[�W���ÐH�\���́A��������B
�@�}�N���t�@�[�W�́A�A���L�h���_����APGE2�APGE1�APGF2���A6-keto-PGF1���iPGI2�̈���ȑ�ӎY���j�A���C�R�g���G���iLT�j�A�g�����{�L�T���iTXB2�j �Ȃǂ��Y������B
�@�v���X�^�O�����W���iPG�j�́A�}�N���t�@�[�W�i�P���j�݂̂Ȃ炸�AT�זE��B�זE��A���̑��A�قƂ�ǂ�����זE����Y�������B
�@
�@PGE2�́A�}�N���t�@�[�W�Ȃǂ����LTB4�̎Y����}�����A�������i�D�����F�������j�̗V����A��������}�����A�����p���iTh2�זE�j�D�ʂ̖Ɖu������U������ƍl������B
�@PGE2���Y�������ƁA���^���̉��ǁi���F�̔^���o��D�����D�ʂ̉��ǁj���A�J�^�����̉��ǁi���t���̕��啨���o�郊���p���D�ʂ̉��ǁj���A�U�������ƁA�l������B
�@PGE2�̎Y�����ߏ�ɍs����ƁATh1�זE�ɂ��זE���Ɖu���ቺ���A���M���Ԃ��������i�M�����������j�A�R�̎Y�����ߏ�ɂȂ�i�A�����M�[�̎��ɂȂ�A�a�C�̉��ɁA�R�̉��̏㏸���������j�B���m�[���_��A���L�h���_�𑽂��ێ悵�Ă���ƁA���̂悤�ȁAPGE2���ߏ�ɎY������̎��iTh2�זE���D�ʂ̑̎��j�ɂȂ��A���ׂ̏Ǐ�┭�M������������A�A�����M�[�������ɂȂ�₷���Ȃ��ƁA�l�������B
�@�Q�l����
�@�E���|�����F�A�g�s�[�̖Ɖu�@���{�����Ȋw��G���@101:1127-1130, 1997�D
�@�E�P�{�m�O�F�T�C�g�J�C���ƃA�����M�[�������|IL-18�ƃA�����M�[�|�@���{��t��G���@��132���E��4��/����17�i2005�j�N2��15���@491�D
�@�E�J�����A���F�W���Ɖu�w�i��Q�ŁA��w���@�A2004�N�j�D
�@�E�R�{��F�A���F�J���[�}���@�x�т̐���\���Ƌ@�\�@�h�u�@���t�E�Ɖu�E������@�i���{�㎖�V��ЁA2002�N�j�D
�@�E�{��M�V�A���F�킩��₷���Ɖu�����@���{��t��G���@���ʍ��i�P�j�@���U����V���[�Y�|�U�V�A2005�N�D�@�E���r�Y���A���F�A�g�s�[���畆�����f���}�E�X�ɂ�����IFN-���Y���ቺ�@����Th1�T�C�g�J�C�����Ì��ʁi���菴�^�j�@���{�����Ȋw��G���@105:354(136), 2001�D
�@�E���i�O�A���F��S�́@�}�N���t�@�[�W�@��g�u���@�Ɖu�Ȋw�R�@�Ɖu�S���זE�A115-149�A��g���X�i1986�N�j�D
�@�E�A���q�A���F�����@TLR�t�@�~���[�ƃE�C���X�����@�E�C���X�@��54���@��2���A145-152�A2004�N�D
�@�E�ؓ������A���R��Y�A���쑾�Y�F�����ۑp�ƃA�����M�[�A�����ǁAVol.36 No.3�A5-6�ŁA�A�X�e���X������Ёi2006�N5���j�D
�@�E���R���q�A���F�����@�H���A�����M�[�f�ÃK�C�h���C���A���{�����Ȋw��G���A110��7���A904-911�A2006�N�D
�@�E��˒�l�A��c����FRS�E�C���X�����ɂ��}���C�ǎx���ƋC�ǎx�b���A�����ȁAVol.41 No.4�A571-578�ŁD
�@�E�]�`�M�F�T���R�C�h�[�V�X�̃A�N�l�ەa�����ƐV���Ȏ��Ð헪�A���{�㎖�V��ANo.4317�i2007�N1��20���j�A68-71�ŁD
�@�E�O�㗝��Y�F�Տ��ƕa���@�P�D�����A�T���R�C�h�[�V�X���߂����āA�Տ���AVol.7 No.8�A1822-1825�Łi1981�N�j�D
�@�E�֓����v�F��109����{�����Ȋw��w�p�W��@����ʃV���|�W�E���F���c���C�ǎx�b�����Â̑�������@�A�����M�[�����ǐ������Ɋւ���`�I�w�i�A�G�s�W�F�l�e�B�b�N����Ɗ����q�A���{�����Ȋw��G���A111��1���A10-15�Łi2007�N�j�D
�@�E�哈�E���F��109����{�����Ȋw��w�p�W��@����ʃV���|�W�E���F���c���C�ǎx�b�����Â̑�������@�A�����M�[�����ǂ̔��Nj@���A���{�����Ȋw��G���A111��1���A16-22�Łi2007�N�j�D
�@�E�������g�A���ш��A�^���N���A����m�G�A�����甿�A�V�c�[�O�A���c��A�r���q�s�F�O��RS�E�C���X�C�ǎx���Ǘ�ւ̃f�L�T���T�]���P��牺���˂̕��y�Ɋւ��錟���A���{�����Ȋw��G���AVol.111 No.2�i��110����{�����Ȋw��w�p�W��^�j�A204�Łi2007�N�j�D
�@�E�⏲�q�A�c���q�F�A���W�I�e���V���ϊ��y�f�A�ŐV�@�Տ�������ABC�A���{��t��G���@��135���E���ʍ��i2�j�A���U����V���[�Y�|70�AS140�ŁA����18�i2006�j�N10���D
�@�E�[�c�r�K�A�R��N�A�k���T���Y�A���c�\��A���㐳�W�A����r�v�F�Ɖu�@�\�ɂ����鈟���̖����A���{�㎖�V��ANo.4343�i2007�N7��21���j�A63-69�ŁD
�@�E�ɓ��T�F���I�����b�A�����V���o�ŁA�Q�O�P�P�N�P�P���R�O����P�����s�D
�@�EM Betz, et al: Prostaglandin E2 inhibits production of Th1 lymphokines but not of Th2 lymphokines. The Journal of Immunology, 1991, 146: 108-113.
�@�EMary F. Lipscomb et al: Dendritic Cells: Immune Regulators in Health and Disease Physiol. Rev. 82: 97-130, 2002.
�@�EFrancesca Alois, et al: CD40-CD154 Interaction and IFN-�� Are Required for IL-12 But Not Prostaglandin E2 Secretion by Microglia During Antigen Presentation to Th1 Cells. The Journal of Immunology, 1999, 162: 1384-1391.
�@�EF Aloisi, et al: IL-12 production by central nervous system microglia is inhibited by astrocytes. The Journal of Immunology, 1997, 159: 1604-1612.
�@�EJe-Jung, et al: The role of PGE2 in the differentiation of dendritic cells: How to dendritic cells influence T-cell polarization and chemokine receptor expression? Stem Cells, 2002, 20:448-459.
�@�ETC van der Pouw Kraan: Prostaglandin-E2 is a potent inhibitor of human interleukin 12 production. Journal of Experimental Medicine, 1995, 181: 775-779.
�@�EMihai G. Netea, et al: From the Th1/Th2 Paradigm towards a Toll-Like Receptor/T-Helper Bias. Antimicrob Agents Chemother, 2005 October; 49(10): 3991-3996.
�@�EEugene Chung, et al: Characterization of Virus-Responsive Plasmacytoid Dendritic Cells in the Rhesus Macaque, Clin Diagn Lab Immunol, 2005 March, 12(3): 426-435.
�ETamir A, et al: Interferon-��2a is sufficient for promoting dendritic cell immunogenicity. Clinical & Experimental Immunology, Volume 142, Number 3, December 2005, pp. 471-480(10).
�@�EEnrico Proietti, et al: Type I IFN as a Natural Adjuvant for a Protective Immune Response: Lessons from the Influenza Vaccine Model, The Journal of Immunology, 2002, 169: 375-383.
�@�b�g�b�v�y�[�W�b�����ƌ����̊W�b�~�j��w�m���b�����w�̒m���b��w�̘b���b�����Ȏ����b�����̕s�v�c�b�����N�W�b